



Comment les plantes supportent les stress alpins ?
PDF
En altitude, les plantes peuvent être soumises à de fortes contraintes : luminosité et rayonnement UV intenses, faible concentration de CO2, grandes variations de température… Autant de paramètres qui varient au cours des saisons mais dont l’amplitude peut aussi fortement changer d’un jour à l’autre comme au cours de la journée. Ces paramètres modulent et parfois perturbent la photosynthèse qui utilise l’énergie apportée par la lumière et permet la croissance et le développement des végétaux vivant dans le milieu alpin. En particulier, l’excès d’énergie lumineuse conduit à la formation de composés toxiques, comme les dérivés réactifs de l’oxygène (en anglais reactive oxygen species, ROS). Pour s’adapter à ces conditions difficiles, les plantes alpines développent diverses stratégies : très petite taille, écran protecteur contre le rayonnement UV, structures anatomiques protectrices, mécanismes pour dissiper l’énergie lumineuse en excès, détoxification des dérivés réactifs de l’oxygène, etc.
1. La végétation dans les étages alpins
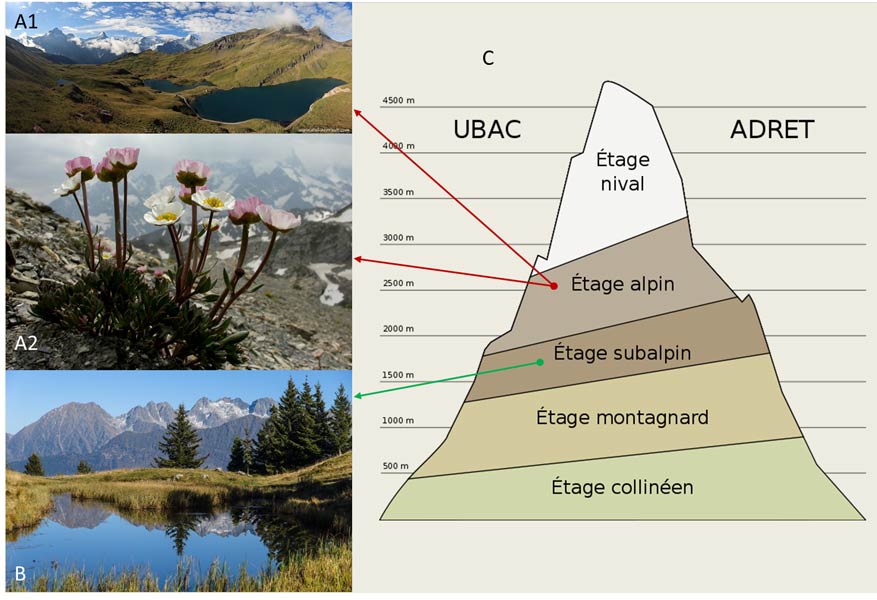
2. Les conditions climatiques et la période de végétation

Ainsi, les plantes pérennes ont l’avantage de néo-former que quelques organes durant la période de végétation pendant laquelle les plantes annuelles doivent former la totalité de leurs organes, ce qui nécessite beaucoup d’énergie pendant un temps limité.

3. La photosynthèse
Durant la période de végétation, les plantes assurent leur croissance et leur développement grâce à la photosynthèse. Dans ce processus, l’énergie nécessaire est apportée par le rayonnement solaire qui permet la synthèse de matière organique à partir du gaz carbonique pris dans l’atmosphère, de l’eau et des éléments minéraux, puisés dans le sol. Parce que la photosynthèse est affectée par de multiples contraintes environnementales comme celles qui sont présentes en altitude [5], il est probable que la maintenance de la photosynthèse sous conditions de stress détermine la survie des plantes à haute altitude. Les principales étapes de la photosynthèse sont résumées dans la figure 4 et détaillées dans [6] (lire « Lumière sur la photosynthèse »). La figure 4 décrit aussi les particularités mises en œuvre par les plantes alpines pour protéger ce processus.
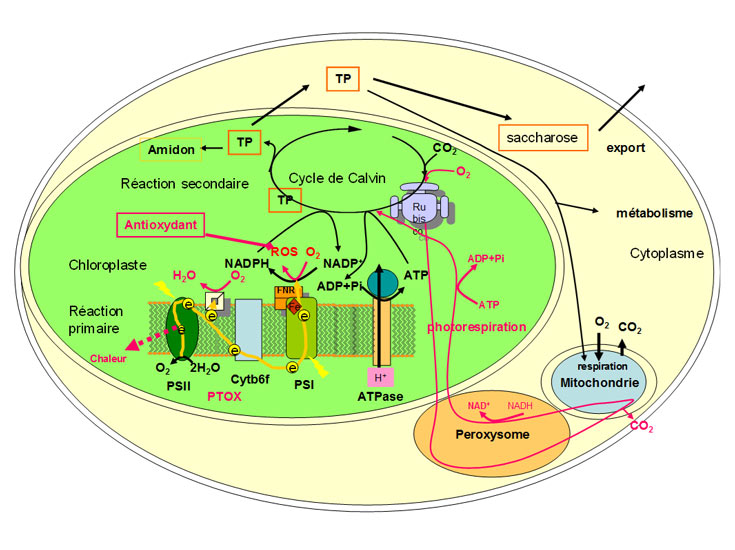
- Une phase primaire se déroule dans la membrane des thylacoïdes : l’énergie solaire est absorbée par des pigments photosynthétiques (chlorophylles formant des antennes collectrices), puis transférée sur deux centres réactionnels (photosystème I et II : PSI et PSII) dont l’activité coordonnée aboutit, via une succession de réactions d’oxydoréduction, à la réduction de NADP+ en NADPHSigle pour la forme réduite du coenzyme Nicotinamide Adénine Dinucléotide Phosphate (NADP). Le NADP est formé à partir du Nicotinamide Adénine Dinucléotide (ou NAD) par liaison d’un phosphate sur le groupe hydroxyle 2′ du ribose associé à l’adénine. Il existe sous une forme oxydée, notée NADP+, et une forme réduite, notée NADPH. Le NADPH est dit porteur de pouvoir réducteur : utilisé dans des catalyses effectuées par des oxydoréductases, il est capable de fournir de l’énergie lors du transfert de leur atome d’Hydrogène, permettant les réactions de réduction nécessaires au fonctionnement cellulaire. par le PSI. Les électrons impliqués proviennent de l’oxydation de l’eau (2H2O → O2 + 4H+ + 4e–) au niveau du PSII. Cette phase primaire induit aussi l’établissement d’un gradient de protons de part et d’autre de la membrane des thylacoïdes qui apporte l’énergie nécessaire pour la synthèse d’ATPAbréviation d’adénosine triphosphate. Nucléoside triphosphate composé d’adénine (base azotée), de ribose (sucre à 5 atomes de carbone) et de trois groupes phosphate formant un groupe triphosphate. Composé à la fois donneur et accumulateur d’énergie présent chez tous les organismes vivants. Sert également de matériaux de construction pour la synthèse des acides nucléiques. (Figure 4).
- La deuxième phase de la photosynthèse se déroule dans le stroma des chloroplastes. Elle consomme l’ATP et le NADPH formés pour fixer le CO2 atmosphérique sur un sucre di-phosphate (activité de la RubiscoAbréviation pour ribulose-1,5-bisphosphate carboxylase/oxygénase. C’est l’enzyme clé permettant la fixation du dioxyde de carbone CO2 dans la biomasse végétale en initiant le cycle de Benson & Calvin, grâce à l’énergie solaire captée par la chlorophylle au cours du processus de photosynthèse.), dont le squelette est formé par 5 atomes de carbone (Ribulose bis phosphate : RuBP) et donner des sucre-phosphates à trois atomes de carbone (Triose-phosphate = TP) (Figure 4). Ce type de photosynthèse qui aboutit rapidement à la formation de composés à 3 carbones est caractéristique des plantes en C3 (C3 pour 3 trois carbones). Ces dernières constituent la grande majorité des plantes alpines.
Les Trioses phosphate (TP), qui contiennent le CO2 assimilé, peuvent être utilisés de plusieurs façons dans les cellules chlorophylliennes : (1) pour régénérer l’accepteur du CO2 ; (2) pour la synthèse de glucose et d’amidon dans le chloroplaste ; (3) pour être exportés vers d’autres compartiments de la cellule où ils fournissent énergie et chainons carbonés pour sa maintenance ; (4) pour la synthèse de saccharose qui est exporté vers d’autres parties de la plante comme source d’énergie et fournisseur de squelettes de carbonés alimentant de nombreuses biosynthèses [6]. Noter aussi que le dioxygène peut se fixer sur le RuBP : cette oxygénation est à l’origine d’une voie métabolique, la photorespiration, qui consomme aussi de l’ATP et du NADPH (Figure 4).
4. Importance des facteurs environnementaux dans la photosynthèse
4.1. Température
Durant la photosynthèse, la température affecte principalement les réactions permettant la fixation de CO2 et d’O2 et la synthèse des sucres, mais aussi les échanges de molécules entre compartiments cellulaires. Les deux principales phases de la photosynthèse ainsi que les processus de transport impliqués sont différemment affectés par la température :
- Les processus biophysiques comme l’absorption de la lumière par les pigments chlorophylliens et la formation de NADPH et d’ATP n’en dépendent pas, ou peu.
- Les réactions biochimiques à l’origine de la fixation de CO2 et d’O2 et de la synthèse des sucres, de même que les échanges de molécules entre compartiments cellulaires en dépendent fortement [7]. En moyenne, une hausse de10°C double la vitesse des réactions biochimiques [6].
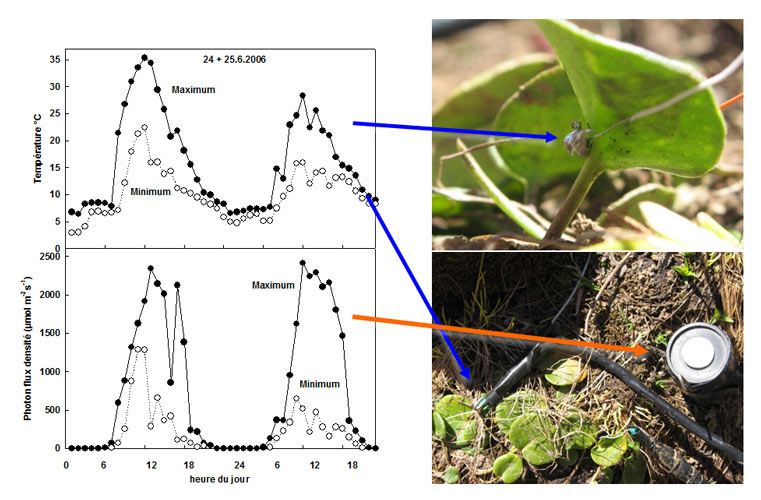
Par exemple, la renoncule des glaciers, sous une lumière apportant 2000 µmoles photons m-2 s-1, fixe environ 15 µmol CO2 m-2s-1 à 23°C (voir Figure 5 pour les ordre de grandeur du flux lumineux), et n’en fixe seulement la moitié lorsqu’elle se trouve à 10°C. Dans le premier cas la lumière n’est pas saturante et la fixation de CO2 peut éliminer l’énergie qu’elle apporte, dans le second par contre, la fixation de CO2 est déjà saturée sous 500 µmoles photons m-2 s-1 et par conséquent la lumière est largement excédentaire [8].
4.2. Lumière
Comme la température, la lumière peut varier soudainement, et ce de plusieurs ordres de grandeur, au cours d’une même journée en conditions alpines (Figure 5). Quand le ciel est couvert, l’intensité lumineuse moyenne n’est pas très différente entre les vallées et les zones situées en altitude [3]. Par contre l’intensité lumineuse maximale par temps clair est beaucoup plus forte à haute altitude, excédant la capacité photosynthétique de la plupart des plantes.
Tout comme une baisse de température, une très forte lumière entraine un excès d’énergie dans les feuilles qui peut endommager les photosystèmes [7]. A cela s’ajoute un rayonnement ultraviolet (UVA et UVB) plus important en altitude [9], susceptible d’impacter directement les structures cellulaires : photosystèmes, mais aussi l’ADN dans le chloroplaste, la mitochondrie et le noyau (lire Impact cellulaire des UV solaires). Au niveau du PSII le manganèse, qui participe à la production d’oxygène (figure 4), et la plastoquinone, qui transporte les électrons entre le PSII et le cytochrome b6/f (cytb6f) absorbent directement les UV et de cette manière leur fonction peut être bloquée [10].
Une plante d’ombre tire parti des intensités lumineuses modérées, fréquentes dans les Alpes, mais elle est stressée si la lumière est trop élevée. A l’inverse, une plante de lumière tolère mieux les fortes intensités lumineuses, mais elle utilise mal la lumière faible [11]. Enfin, des plantes alpines de pleine lumière comme Soldanella alpina, Homogyne alpina ou Ranunculus glacialis (Figure 2) ne montrent pas d’adaptation spécifique à la lumière faible ou forte [11].
4.3. Autres facteurs
D’autres facteurs environnementaux alpins affectent l’activité photosynthétique :
- La diminution de la pression de l’air en altitude, par exemple, réduit la disponibilité du CO2 [7].
- Le vent -dont la force dépend de la topographie- augmente le risque de dessèchement des plantes.
- En favorisant le ruissèlement, la pente diminue aussi la rétention de l’eau par le sol [3].
Or, en cas de stress hydrique, une plante réagit en fermant ses stomates (cellules spécialisées contrôlant les échanges gazeux de la feuille) : cela limite l’apport de CO2 et freine la photosynthèse. Ici encore, les réactions biochimiques de la photosynthèse ne permettent pas de résorber l’excès d’énergie lumineuse reçue par la plante.
En résumant, les facteurs climatiques (tels que lumière, température, humidité) varient fortement, rapidement et avec de grandes amplitudes dans les montagnes. Si l’on expose la majorité des plantes de plaine à de telles variations, elles ne survivent pas, parce que la photosynthèse n’est pas capable d’utiliser le surplus d’énergie [7]. Ce surplus d’énergie peut être transféré à l’oxygène en formant des espèces moléculaires réactives de l’oxygène (ROS) (Figure 4 et [12]). Les ROS sont des molécules potentiellement très destructrices ; elles peuvent endommager non seulement l’appareil photosynthétique et particulièrement le photosystème II (photoinhibitionProcessus par lequel un excès de lumière diminue la vitesse de la photosynthèse chez les organismes capables de l’effectuer.), mais aussi l’ensemble de la cellule (lire La vie fixée des plantes). Au final, la capacité photosynthétique de la plante risque d’être inhibée par les ROS, et sa capacité à réparer les dommages fortement diminuée [11].
4.4. Quelles contraintes pour la photosynthèse des plantes alpines ?
L’illustration des contraintes imposées par l’environnement alpin peut être donnée, par exemple, dans deux cas extrêmes.
Certaines plantes alpines comme Soldanella alpina, Geum montanum et Homogyne alpina gardent quelques feuilles vertes durant l’hiver. La photosynthèse de ces feuilles démarre aussitôt après la fonte des neiges, dans des conditions alliant les fortes intensités lumineuses à d’importants gradients thermiques [7]. Dès le déneigement, ces espèces alpines doivent mettre à profit cette courte période pour leur photosynthèse, parce qu’elles seront recouvertes par les autres végétaux quelques semaines plus tard. Une inactivation de la photosynthèse sous climat extrême peut être létale pour ces plantes.
Mais la majorité des plantes alpines ne gardent pas leurs feuilles en hiver et doivent mobiliser rapidement leurs réserves pour former leurs premières feuilles et assurer leur cycle de développement durant une courte période favorable. Le cas de Ranunculus glacialis (voir Figure 2), qu’on ne trouve qu’au-dessus de 2200 m, montre que cela peut être encore plus complexe. En effet chez cette plante, on calcule que les réserves mobilisées pour la formation des feuilles représentent environ 30 jours de la photosynthèse ce qui peut à peine être réalisé durant la saison de croissance [3]. Comme les fleurs de Ranunculus glacialis ont besoin de deux années pour leur maturation [12], cela suggère que l’énergie reçue durant une seule période de végétation n’est effectivement pas suffisante pour terminer son cycle de développement en un an. Ranunculus glacialis ne peut donc pas risquer une inactivation de la photosynthèse. Le nanisme de plusieurs espèces alpines peut être aussi considéré comme une stratégie limitant l’investissement d’énergie dans la formation de tissus photosynthétiques.
5. Acclimatations chez les plantes alpines
L’acclimatation met en jeu des réponses pour minimiser l’amplitude des variations des paramètres physiques de l’environnement au niveau de la plante, avant qu’ils n’exercent leurs effets négatifs au niveau cellulaire et moléculaire. Lorsque ces réponses sont insuffisantes, l’acclimatation se manifeste par le biais d’autres protections visant à éviter les destructions des cellules ou de ses organites.
Les plantes alpines ont souvent une architecture particulière en coussin (voir Figure 3), en rosette ou en tussock (lire Héritage ou convergence). Ce type de morphologie met les plantes dans un microclimat favorable [3] avec : (1) une diminution de l’intensité lumineuse et des UV reçus par les tissus foliaires ; (2) une augmentation de l’humidité au niveau des feuilles avec protection contre le vent, limitant le dessèchement ; (3) des variations plus faibles des températures que dans l’air environnant. L’acquisition de ce type de morphologie représente une première réponse d’évitement. Bien sûr, la croissance de ces plantes est bien plus lente que celle des plantes qui ne forment pas ces structures : c’est le prix à payer.
Comme décrit plus haut, les variations thermiques de très grande amplitude conjuguées à une forte luminosité peuvent induire la photoinhibition du photosystème II et la formation des ROS.
Comparées aux plantes de plaine, les plantes alpines que nous avons étudiées [7] résistent beaucoup mieux à la photoinhibition. Pour y parvenir, elles utilisent différents mécanismes de protection dont la plupart existent aussi chez les plantes de plaine. Ce sont bien des mécanismes d’évitement se manifestant à plusieurs niveaux d’organisation (pour les détails de plusieurs mécanismes de protection voir la figure 4). Ils peuvent se diviser en deux catégories. Des mécanismes « en amont » qui évitent l’arrivée trop rapide de l’énergie sur les centres réactionnels et des mécanismes « en aval » qui maintiennent l’utilisation des électrons produits (c’est-à-dire in fine l’utilisation de l’énergie lumineuse captée par les antennes) même dans des conditions adverses (voir Figure 6).
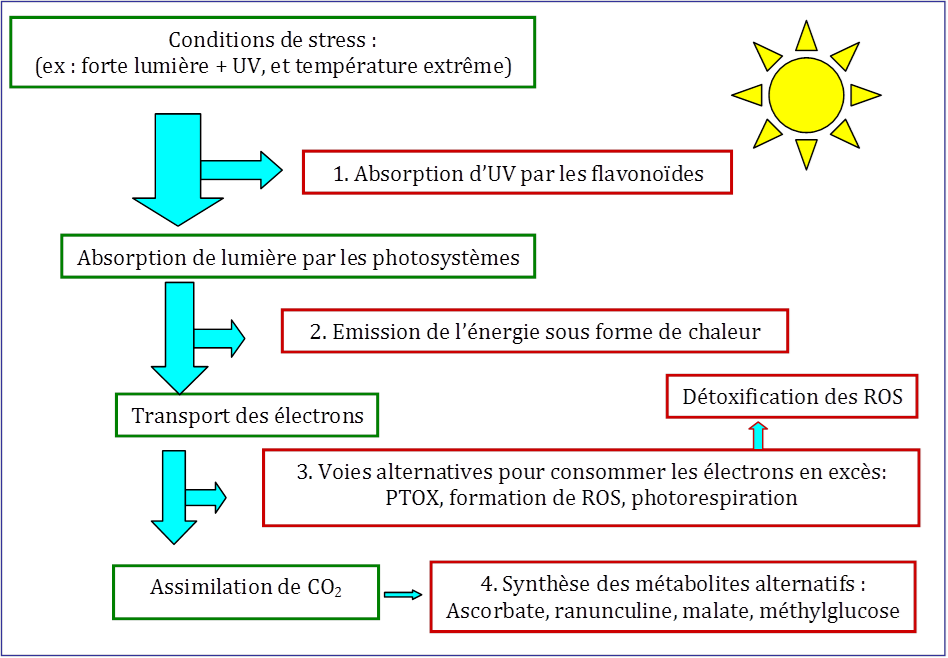
- Protection contre les UV. Au niveau de l’épiderme, quelques espèces synthétisent des flavonoïdesMétabolites secondaires des plantes partageant tous une même structure de base. Avec plusieurs milliers composés, les flavonoïdes constituent la plus importante catégorie de polyphénols et représentent une gigantesque famille d’antioxydants. Les flavonoïdes sont à l’origine des teintes brunes, rouges et bleues des fleurs et des fruits., pigments qui absorbent le rayonnement UV très énergétique. Ainsi, les UV n’excitent pas les cellules sous-jacentes. Cet écran de flavonoïdes protège ainsi les photosystèmes et empêche la destruction de l’ADN des parenchymes chlorophylliensTissus constitués de cellules chlorophylliennes comportant de nombreux chloroplastes. Siège de la photosynthèse, ils participent aux fonctions de nutrition. Ils représentent une partie importante de l’intérieur des feuilles.. Chez Soldanella alpina, par exemple, une protection de cette nature, induite par la lumière forte, augmente avec l’altitude [13].
- Réémission de l’excédent d’énergie absorbée. Les antennes collectrices de lumière changent de conformation chez les plantes éclairées ; parallèlement, une partie des chlorophylles excitées passent leur énergie à des caroténoïdes (la zéaxanthine par exemple). Cela permet à une partie de l’énergie lumineuse d’être dissipée sous forme de chaleur (Figure 4). De cette façon, la quantité d’énergie arrivant sur les centres réactionnels est contrôlée. Sous lumière excédentaire, la fraction lumineuse dissipée thermiquement est plus élevée que celle qui se dirige vers les centres réactionnels. La dissipation thermique est un moyen rapide de faire disparaitre les états excités de la chlorophylle. Cependant, si chez quelques espèces alpines, comme Soldanella alpina et Geum montanum, ce mécanisme d’évitement est très efficace, chez d’autres comme Ranunculus glacialis il s’avère peu opérationnel et d’autres mécanismes sont activés (voir plus bas) [7].
- Transfert d’énergie sur des accepteurs alternatifs via un flux d’électrons. Dans le chloroplaste, l’oxydase alternative (appelée PTOXEnzyme des membranes des thylacoïdes des chloroplastes, qui catalyse l’oxydation du pool de plastoquinone, d’où son nom d’oxydase alternative (PTOX).), branchée entre les deux photosystèmes (Figure 4), accepte les électrons fournis par le PSII et les transfère sur l’oxygène moléculaire ce qui crée de l’eau. Cette enzyme peut diminuer ainsi la pression électronique et donc la formation des ROS sous lumière excédentaire. Par comparaison avec les plantes de plaine, le contenu des plantes alpines en PTOX est élevé et sa concentration augmente avec l’altitude chez Geum montanum [7], [8].
- Maintien du métabolisme photosynthétique à basse température. Si l’apport en CO2 limite la synthèse des sucres dans la deuxième phase de la photosynthèse, la Rubisco catalyse la fixation de l’oxygène moléculaire sur le RuBP. C’est la première étape de la photorespiration (Figure 4). La photorespiration consomme de l’énergie, sous forme (a) de carbone (RuBP), (b) d’électrons produits pendant la première phase de la photosynthèse et (c) d’ATP. L’activité de la photorespiration, habituellement négligeable à basse température, devient importante lorsque cette dernière dépasse les 20°C. Chez Ranunculus glacialis qui ne supporte pas la chaleur, la photorespiration est déjà active quand la température est fraîche [7], [8].
- Synthèse de métabolites particuliers. Certaines plantes alpines synthétisent des métabolites généralement absents ou présents en faible quantité chez les plantes de plaine. La fonction de ces métabolites n’est pas souvent comprise. Mais leur synthèse consomme une partie de l’énergie captée par l’appareil photosynthétique et peut par, ce biais, aider à sa protection. Ainsi, Ranunculus glacialis accumule des quantités importantes de ranunculineGlucoside produite par les plantes comme les renoncules (Ranunculaceae). Molécule très instable, elle est hydrolysée en une lactone irritante : la proto-anémonine. et de malateSel de l’acide malique, un acide dicarboxylique très répandu dans le règne végétal et naturellement présent dans les fruits, dont il contribue à la saveur agréable. Le malate est un intermédiaire du cycle de Krebs, l’une des voies métaboliques majeures de respiration cellulaire chez presque tous les êtres vivants, et intervient dans le cycle de Benson & Calvin, qui entre dans le cadre de la photosynthèse. Utilisé comme additif alimentaire, sous le numéro E296. [14]. Geum montanum contient des concentrations élevées de méthylglucoseMonosaccharide (glucose) portant un groupe méthyl (CH3)., alors que Soldanella alpina renferme dans ses feuilles des quantités record d’ascorbate (vitamine C) [15]. L’ascorbate fait partie du système antioxydant qui protège la plante vis-à-vis des ROS.
- Détoxification des formes très réactives de l’oxygène (ROS). Les contraintes environnementales augmentent la formation de ROS, en particulier au cours des premières étapes de la photosynthèse. Dans la Figure 4, la formation de ROS est indiquée au niveau du PSI : c’est la voie majeure de la formation des ROS à la lumière. Pour dégrader les ROS en oxygène, les plantes utilisent un ensemble de réactions enzymatiques avec les métabolites associés. Dans les feuilles de Soldanella alpina, le système antioxydant s’appuie sur la présence de vitamine C et de vitamine E, ainsi que sur des enzymes impliquées dans des processus d’oxydoréduction [7]. Chez Potentilla saundersiana, la part du système antioxydant dans la résorption de l’excès d’énergie emmagasinée dans les feuilles augmente avec l’altitude [16].
En résumé, c’est tout un ensemble de mécanismes de protection qui permettent aux plantes alpines non seulement de survivre, mais aussi de se développer dans un habitat de plus en plus contraignant avec l’altitude. Ces mécanismes se complètent et s’additionnent pour faciliter la vie des plantes alpines (Figure 6).
Références et notes
Photo de couverture : Linaire des Alpes (Linaria alpina) dans les schistes du Galibier (2600 m). © S. Aubert/SAJF
[1] Ozenda P., La végétation de la chaîne alpine dans l’espace montagnard européen, Masson, 1985
[2] Fischesser B., La vie de la montagne, Éditions de La Martinière, Paris, 1998
[3] Körner C., Alpine plant life, Springer Verlag, Berlin Heidelberg, 1999
[4] Larcher W., Kainmüller C., Wagner J., Survival types of high mountain plants under extreme temperatures, Flora, 205, 3-18, 2010
[5] Lütz C., Plants in Alpine Regions, Cell Physiology of Adaptation and Survival Strategies, Springer, Wien New York, 75-97, 2012
[6] Raven P., Evert R., Eichhorn S., Biology of Plants, Sixth Edition, W.H. Freemann and Company/Worth Publishers, 1986
[7] Streb P., Cornic G., Photosynthesis and Antioxidative Protection in Alpine Herbs, In Lütz C. (Ed) Plants in Alpine Regions: Cell Physiology of Adaptation and Survival Strategies, Springer Wien New York, pp 75-97, 2012
[8] Streb P., Josse E.-M., Gallouët E., Baptist F., Kuntz M., Cornic G. (2005) Evidence for alternative electron sinks to photosynthetic carbon assimilation in the high mountain plant species Ranunculus glacialis. Plant, Cell Environment 28, 1123-1135)
[9] Barry R., Mountain weather and climate, Cambridge University Press, Third Edition, 2008
[10] Teramura A., Ziska L., Ultraviolet-B radiation and photosynthesis, In “Advances in Photosynthesis Vol. 5: Photosynthesis and the Environment”. Ed. N.R. Baker, pp 435-450, Kluwer Academic Publishers, Dordrecht,1996
[11] Walters R., Towards an understanding of photosynthetic acclimation, Journal of Experimental Botany 56, 435-447, 2005
[12] Ort D., Baker N., A photoprotective role of O2 as an alternative electron sink in photosynthesis? Current Opinion in Plant Biology 5, 193-198, 2002
[13] Laureau C., Meyer S., Baudin X., Huignard C., Streb P. (2015) In vivo epidermal UV-A absorbance is induced by sunlight and protects Soldanella alpina leaves from photoinhibition. Functional Plant Biology, 42, 599-608
[14] Streb P., Aubert S., Gout E., Bligny R. (2003) Reversibility of cold- and light-stress tolerance and accompanying changes of metabolite and antioxidant levels in the two high mountain plant species Soldanella alpina and Ranunculus glacialis. J. Exp. Bot. 54: 405-418
[15] Bligny R., Aubert S., Specifities of metabolite profiles in alpine plants. In Lütz C. (Ed) Plants in Alpine Regions: Cell Physiology of Adaptation and Survival Strategies, Springer, Wien & New-York, pp 99-120, 2012
[16] Lan Ma, Xudong Sun, Xiangxiang Kong, Jose Valero Galvan, Xiong Li, Shihai Yang, Yunqiang Yang, Yongping Yang, Xiangyang Hu, Physiological, biochemical and proteomics analysis reveals the adaptation strategies of the alpine plant Potentilla saundersiana at altitude gradient of the Northwestern Tibetan Plateau, Journal of Proteomics, 112, 63-82, 2015
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Pour citer cet article : STREB Peter, CORNIC Gabriel, BLIGNY Richard (27 juin 2017), Comment les plantes supportent les stress alpins ?, Encyclopédie de l’Environnement. Consulté le 20 avril 2024 [en ligne ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/vivant/plantes-supportent-stress-alpins/.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.