





地衣,令人惊叹的开拓者

身为出色的开拓者,地衣征服了最极端的环境。无论是高山之巅的磐石山岩,还是浪花冲刷的岩石海岸;无论是鲜少冷却的熔岩流,还是热带森林的树木枝头;无论是房顶的瓦片,还是老楼的石墙,都可以任它们生长!全球有近20000种地衣,这一数量充分彰显了其丰富的生物多样性。地衣不仅形状和颜色千变万化,而且具备对极端条件的耐受力,这些特质始终令科学家们为之着迷。
1. 地衣,多姿多彩的风景线

图1. 一些常见的地衣。图片内容包括:名称(附照片)、地衣体的特征、地衣体与基质之间的关系。
[图源:© J. Asta]
地衣通常不太起眼,但在生长旺盛的环境,如高海拔地区或沿海地区,它们往往能够呈现出引人注目的缤纷色彩。从黑色到白色,再到各种色调的蓝、绿、黄、橙或红色,地衣的颜色像调色盘一样丰富多变。
地衣的形态同样千变万化,在植物界可谓无出其右。与高等植物不同,地衣没有根、茎和叶子。它们的营养器官被称为地衣体(thallus),具有形状复杂多样的特征,能够以各种方式固定在多种多样的基质(substrate)上(图1)。
部分地衣体呈现叶状或鳞片状,有一些则为直立或倒悬的枝状,还有一些地衣体是壳状的,紧附于基质生长。
地衣体或牢固或松散地附着在基质上。基质的种类极为丰富,包括树干和树枝、岩石和崖壁(石灰岩或硅岩)、古迹、土壤、路边等等。
不同种类的地衣和基质之间的接触面积相差悬殊,可以大到几乎覆盖整个地衣体,也可以小到仅有一个点。
2. 地衣共生:一种原始的组合
2.1. 地衣中存在怎样的共生关系?
根据国际地衣学协会(International Association for Lichenology)的定义,“地衣是真菌和光合生物的复合共生体,是具有特定结构的、稳定的植物”。
科学家首先对共生生物分别进行培养和研究,然后将两种共生生物合并培养,从而获得共生搭档的生理信息和彼此间营养交换的信息。
真菌以名为假根(rhizinae)的小丝为根系,将地衣固定在基质上,并以其高生物量保护光合生物。此外,真菌作为异养生物,还能够为光合共生生物提供水、矿物盐和某些维生素(如维生素C)[1]。
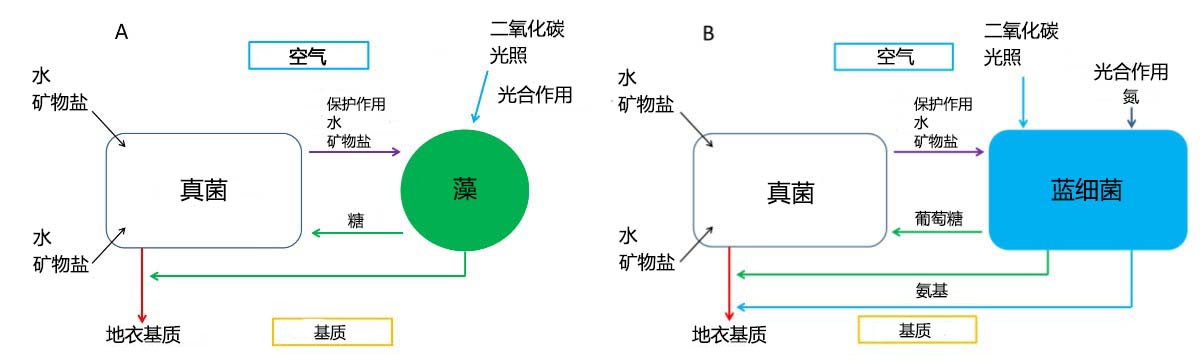
光合共生生物*是碳基自养生物,能够进行光合作用,为真菌提供含碳物质。绿藻的产物还包括糖的衍生物多元醇(polyols)[2],以及维生素B(图2A)。
在以蓝藻代替藻类的共生体中,蓝藻能够生成葡萄糖(glucose)(图2B),真菌则将多元醇和葡萄糖转化为甘露醇(manitol)*和阿拉伯醇(arabitol)*。除此之外,蓝藻还有固氮的作用,可以将大气中的氮以铵的形式传递给真菌(参见《靠空气生存的植物》)。
真菌和藻类的结合非常新颖:真菌产生大量的次生代谢产物,即地衣成分(lichen substance)或地衣酸(lichen acid),这类产物在仅有真菌的情况下是不会产生的。目前已发现超过700种地衣代谢物[3],这些化合物赋予地衣多种特定属性,包括将自身固着于基质、维持水分平衡、调节光合作用、以及防止光辐射或温变对自身产生影响。这些次生代谢产物使地衣能够在不含有机物的基质(如石头、岩石、熔岩等)上生长。
上述产物也实现了地衣在制香[4]、制药和医疗(详见焦点:《地衣在治疗方面的应用》)、染料制造[5]等领域的多重应用。
2.2. 令人惊讶的发现

目前发现与肺衣(Lobaria pulmonaria)[6](图3)有关的细菌已超过800种,这种地衣常在药典中出现,并因其药用价值而闻名(详见焦点:《地衣在治疗方面的应用》)。这些细菌能够参与实现多种功能[7],如供应氮、磷和硫,防御病原体,抵抗特定非生物因素(温度、湿度等)的危害,抵抗重金属危害,通过向藻类提供维生素B12协助光合作用,合成藻类和真菌的生长激素(即吲哚乙酸),降解地衣体的老化部分等。
地衣的共生真菌一般是多细胞的。因此,在发现许多地衣物种,特别是梅衣科(Parmeliaceae)地衣物种属于由特定单细胞担子菌(unicellular basidiomycetes)*组成的另外一种真菌共生体时,地衣学界可谓一石激起千层浪[8]。这些单细胞担子菌参与了地衣体结构的生长发育和地衣次生代谢产物的生产过程。
因此,地衣共生不只是两个或三个组分之间的伙伴关系,而是相当于一个真正的复杂生态系统(详见《共生与寄生》)。因为共生体具有其非共生亲本所没有的组成特征,地衣共生扩大了生物多样性,包括系统多样性(全世界共有超过20000种地衣)、形态多样性(地衣作为共生体,能够创造单独的真菌和光合生物所不具备的新形态)和生化多样性(地衣能够合成特有的次生代谢产物)。
2.3. 共生的结果
共生对地衣适应极端条件的能力、寿命和特殊的生长模式有着显著的影响。事实上,地衣的生长通常极为缓慢,每年仅生长0.1到1毫米不等[9]。其中,壳状地衣生长最慢。
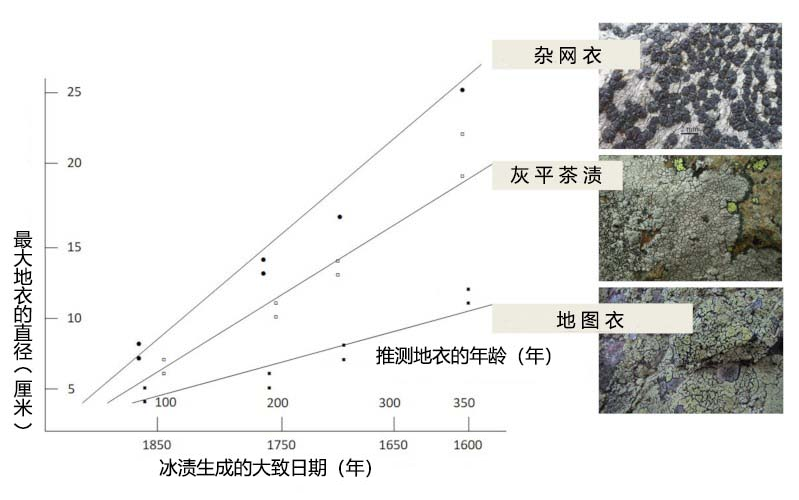
对壳状地衣而言,直径仅数十厘米的地衣体便已意味着数百年的高寿。地衣测年法(lichenometry)以地衣的低生长速率和漫长寿命为基础,通过已知的地衣生长速率和最大地衣体尺寸的测量数据来评估地衣的年龄,进而确定基质[10]的年龄。在对地图衣(Rhizocarpon geographicum)的研究中,对地衣体的测量结果显示,其形成于300-400年前,通过该结果,可推断出冰川冰碛的年龄[10](图4)。
3. 地衣及其生态
从海边岩石到高山之巅,从干旱的沙漠到高纬度地区,除了深海、污染严重的地区和动物活体组织,地衣几乎能在所有的环境中生长。任何物种的生存都需要特定的生态条件,主要包括物质供应类型、气候因素和生物间关系。
3.1. 基质与生境的类型

地衣的形态不同,对基质的依赖程度也不同:壳状(crustose)地衣对基质依赖性非常强,叶状(foliiose)地衣和枝状(fruticose)地衣对基质的依赖性则较弱。地衣可生长于树木(corticolous lichens,树生地衣),土壤(terricolous lichens,陆生地衣),岩石(saxicolous lichens,岩生地衣),苔藓,旧木材,常绿叶片上。有些地衣甚至可在其他地衣上寄生。各种各样的物体表面都可以成为地衣的基质,如旧墙、木制或金属栅栏(柱子)、铁轨、旧皮革、塑料、玻璃等(图5)。
地衣会根据基质调节水分情况以及地衣体的固定方式,主要影响因素包括机械和物理因素(如组织结构和树皮孔隙度,岩石密度和非均质性,稳定性,结构,粒度和土壤孔隙度等)及化学因素(如pH值、基质的矿物化学组成或有机成分等)。
有些地衣的存在表明其基质中存在碳酸钙,这类地衣被称为喜钙物种(calcicolous species)。有些地衣则是嗜酸物种(acidophilous species),会寻找酸性更强的土壤、岩石或树皮作为基质。有些地衣可作为金属元素指示物,它们或只在含有某类金属的基质上生长,或对土壤和岩石中的某类金属具有一定的耐受性。如果某块岩石上覆盖有铁锈色的地衣体,即可确认其为含铁矿石。通常来说,鸟类粪便积累会使基质富含以氮为首的可溶性矿物质,这类基质也有利于特定地衣的生长,丽石黄衣(Xanthoria elegans)便是其中一种。
3.2. 气候因素的影响
大气能够向地衣提供其所需的水、二氧化碳和矿物质,是非常重要的影响因素。
3.2.1. 水
水在地衣生态中起着至关重要的作用,因为地衣体的水化程度决定了地衣体的基本功能。有些地衣可以从湿润时的活跃状态快速调节到干燥时的缓慢状态,反之亦然。地衣随环境湿度增加而恢复活跃状态的现象被称为复生(reviviscence phenomenon)。在干燥状态下,地衣的含水量约占自身干重的15-20%,而在湿润状态下,该比例可高达200-350%。胶状地衣(Gelatinous thallus lichens)富含粘液[11],含水量最高,可达其干重的3500%。地衣用液态水浸泡将近1-2分钟便可达到湿润状态,但对蒸汽的吸收速率较慢,有时需要几周时间才能达到湿润状态。而干燥过程则与之相反,发生得非常迅速。

在干燥状态下,地衣的气体交换活动几乎微不可察。然而,一旦湿润,地衣的呼吸作用和光合作用很快就会恢复。对高海拔地区丽石黄衣(Xanthoria elegans)[12](图6)的研究表明,地衣水化后,呼吸作用仅需一秒即可恢复,光合作用的恢复也仅需一分钟。呼吸作用之所以恢复得如此之快,是因为地衣在脱水过程中积累了6-P葡萄糖酸盐,同时保存了核苷酸。在丽石黄衣的两种共生体中,高浓度的多元醇(详见参考资料及说明[2])有助于在干燥状态下保护细胞组分,维持细胞内结构的完整性。水化发生后,6-P葡萄糖酸盐立即代谢,为呼吸作用提供必要的底物。在低温条件下,丽石黄衣的湿地衣体同样具有很强的光合作用活性,这一特质有助于其利用高海拔地区的融雪等机会,短暂地进行水化,从而在高山地区的恶劣气候条件下实现生长。
这也解释了为什么即使身处日照充足的环境,生长于高海拔地区岩石基质上的地衣也只会在露珠初现的早晨或雨后几小时内进行光合作用。这样的高抗性使得丽石黄衣(Xanthoria elegans)得以分布于喜马拉雅山脉海拔7000多米处,成功位列能在最高海拔生长的物种。
许多地衣生长在地面的小石头上,或者离地面很近的小卵石上,它们生活的气候条件与那些以广袤大地为基质的地衣截然不同——不仅热量和水分波动更大,而且有晨露提供额外的水分供给。有些地衣只栖息在悬垂的墙壁上,因而总能免受雨水侵袭。相反,另一些地衣则可以承受暂时性的雨水冲刷,甚至在淡水或海水中生长。因此,在岩石海岸,从巨石的顶部到底部的藻类,我们可以自顶向下观察到典型的地衣植被分层(详见《岩石海岸的生物多样性:分区和生态关系》)。
一些地衣对大气中的盐分具备一定的耐受性,因而能够在浪溅区定居。智利海岸的一些海洋地衣表面覆盖有一层盐壳,能够起到吸湿的作用,将地衣的吸水率提高至原来的三倍。
3.2.2. 光照和温度

光照能够增强光合作用,在一定的光强下,光合作用的强度可能超过呼吸作用。有些地衣偏爱光照环境,甚至能够承受高强度的阳光。另一方面,有些地衣则生长在较阴凉的地方,例如林荫下。而对于岩生地衣而言,背光悬垂的岩石表面就是适宜其生长的不二选择。

在干燥状态下,地衣对极端温度具有极强的抗性。在高海拔地区,地衣可以承受长时间的极低温度或极高温度,例如,太阳辐射可导致岩石表面温度飙升至50°C,但地衣仍可在这样的基质上存活。然而,全球范围内的长期温度变化正在改变地衣的整体分布,目前,地衣的栖息地范围正随全球变暖向北扩展,卷叶皱衣(Flavoparmelia soredians)(图7)就是其中一员(这种迁徙特性常见于鸟类种群,详见《鸟类如何适应气候变化?》)。
在高海拔地区,科研人员开展了夏季紫外线辐射对冰岛地衣(Cetraria islandica)(图8)的影响研究 [13]。研究结果表明,辐射对光合色素和夜间测量的呼吸作用影响不显著。另一方面,中波紫外线(UVBs)导致12种酚类化合物中的8种浓度显著降低。这意味着冰岛地衣体内的酚类化合物具有相当强的抗紫外线能力,也解释了为什么这种地衣能够承受高海拔地区的辐射。
3.2.3. 风的作用
风有两种作用方式:
一是间接的生理作用,提升脱水速度。
二是直接的机械作用,在地衣体碎片和营养繁殖体的传播中扮演重要角色。
一个非常特别的例子是,分布于沙漠到半沙漠草原上的一些叶状地衣只会在湿润状态下进行光合作用,并将上皮层暴露于阳光下,处于干燥状态时则会把自己卷起来。在这种形态下,它们很容易被风携带并传播,这种地衣因此也被称为“不稳定(erratic)”地衣。
在模拟太空条件(真空、高紫外线辐射)下进行的实验表明,各种地衣的孢子都具有极强的抗性,尤其是丽石黄衣的孢子。[14]
3.3. 与其他生物的关系
3.3.1. 地衣彼此间及地衣与植物间的关系

在树皮或岩石上,壳状地衣通常是第一批定居者,叶状地衣和枝状地衣则紧随其后。如果生态条件合适,苔藓植物(bryophytes)和维管植物(vascular plants)也常会后来居上(图9)。土壤上的地衣在上层植被过于密集的地方无法生存,因为它们体型过小,容易受高等植物的影子和落叶阻碍。
3.3.2. 动物与人类的作用
动物和人类会踩碎地面上的地衣体,也会增加地表铵盐、硝酸盐等物质的含量,改变环境的化学性质。此外,人类自身还扮演着一个特殊的角色:
人类能够创造新的基质(墙壁、砂浆、水泥、瓷砖、矿石截面等),使特定的地衣植物可以在上面定居;
人类破坏气候(如减少大气水分,提高环境温度,产生灰尘或焦油污染等),削减有利于地衣栖息的环境(如砍伐活动造成森林面积减少,城市、荒野、干旱草原的扩张毁坏灌木和树林等)。
对于曾生长过苔藓植物和地衣类群的岩石板块,地衣大约需要一个世纪以上的时间才能重新定殖。所幸目前移植技术已经可以实现修复,无论是重建已经灭绝的地衣群落(如肺衣[15]),还是在伐木前进行预防性重建。
4. 地衣,开拓者?
4.1. 地衣如何在新的基质上定居?
地衣具备适应极端条件的能力,可以在营养匮乏的基质上生存,因而被视为真正的开拓者。
地衣孢子的扩散是定殖的第一步(详见焦点:《地衣:共生生物》)。地衣的孢子一旦成熟,就会被猛烈喷出子囊,而后落在基质上。孢子能够快速生长,产生分支的菌丝。为了形成新地衣,菌丝必须要有一个藻类搭档。菌丝和藻类之间的识别通常是由凝集素(lectins)*这类分子渗透细胞壁来实现的。随后,二者形成一个未分化的地衣幼体,并逐渐生长为成体,大多数时候还会长出生殖器官。
4.1.1. 岩石定植
地衣是通过物理和化学作用定殖在岩石上的。在石灰岩中,地衣产生的草酸(oxalic acid)能够促进碳酸钙溶解并转化为草酸钙。在酸性岩石中,地衣分泌物可以通过物理作用分解岩石矿物质。
一项关于地衣对历史遗迹作用的研究表明,地衣体菌丝在花岗岩基质中渗透的深度可达2毫米。[16] 地衣体能够真正地“消化”并吸收二氧化硅。
地衣也能迅速在火山喷发时喷出的熔岩上实现定殖。这就不得不提到多型珊瑚枝(Stereocaulon vesuvianum),这种含有绿藻和蓝藻的地衣首次发现于维苏威火山的沉积物中,其中一种名为Dyctionema pavonia,生长于法属瓜德罗普岛的苏菲埃火山岩壁上。
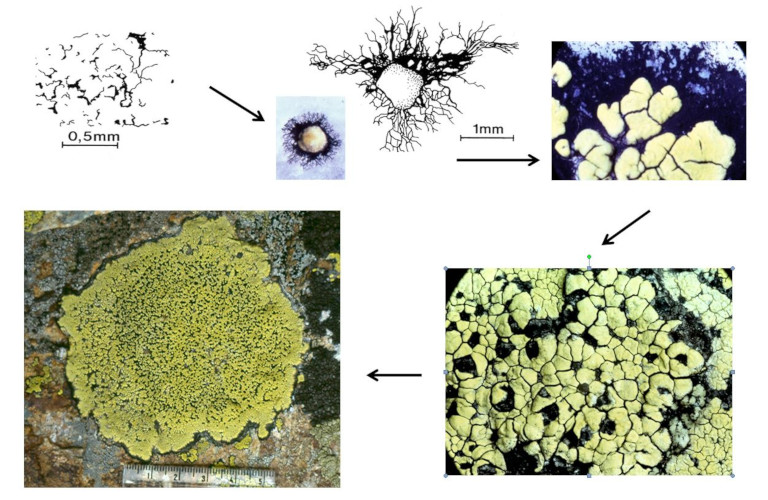
对高山环境中石英岩(quartzite)*的定殖研究有助于我们理解地图衣(Rhizocarpon geographicum)的定殖策略(图10)[17]。这种壳状地衣体由包含真菌和藻类的隔间(或网隙)组成。真菌孢子在萌发时会形成黑色的放射状菌丝体(基质层),并在其基础上形成初始的网隙。多个网隙在菌丝体上汇合,形成首个地衣体。该地衣体又与其他地衣体汇合,最终形成的完整地衣体直径可达15厘米[16]。
4.1.2. 土壤定植

羊角衣属(Baeomyces)和石蕊属(Cladonia)地衣是真正的土壤开拓者。山区的观察结果表明,在用于建造滑雪坡道的裸露山坡上,最初有两种生物生长:一是地衣,更确切地说是淡红羊角衣(Baeomyces rufus);二是苔藓,更确切地说是疣小金发藓(Pogonatum urnigerum)。羊角衣能够开拓裸露的基质表面,形成一种菌丝毡,这种菌丝毡非常重要,能够帮助稳定表层土壤(图11)[18]。疣小金发藓则采取不同策略,比地衣扎根更深。二者共同构成了真正的地衣-苔藓复合体,有利于更复杂的土壤的形成[18]。
石蕊属地衣柱体(podetions)*的基部能够分解并释放菌丝,这些菌丝可逐渐融入泥土颗粒中,与之共同构成一个真正的地衣-土壤复合体,这种复合体通常富含细菌菌落,且同前文的地衣-苔藓复合体一样,有利于高等植物随后定殖。[19]
在地球上的某些区域,地衣是生态系统的重要组成部分。虽然全球范围内,地衣区系(lichenic flora)和显花植物区系(phanerogamic flora)*的平均分布比重从1:10到1:1不等,但在北极地区,二者的比率可能超过100:1。该地遍布苔原和温带森林,在这些地貌下,以鹿蕊(Cladonia rangiferina),即麋鹿地衣为首的几种地衣是麋鹿、驯鹿、驼鹿、欧洲盘羊等多种野生动物的常见食物来源,一些家养动物偶尔也会以地衣为食(详见焦点:《地衣与动物的交互》)。
在干旱和半干旱地区,地衣有助于形成沙漠土壤上的“生物结皮”(图12)。地衣、蓝藻、真菌、苔藓植物和藻类等多种生物都可参与构成生物结皮,在土壤表面的上几毫米处紧密共生。全球约有5000万平方公里的土地为这类结皮所覆盖,相当于5个欧洲的大小。生物结皮在保护土壤、增强肥力、防止水土流失和荒漠化等方面发挥着重要作用。据估计,仅结皮就可固定7%的二氧化碳,与人类每年燃烧化石燃料的排放量相当。[20]

4.2. 地衣与定殖的起源
和其他诸多涉及生命的学科一样,地衣学也面临着起源的问题。大约50年前,人们仍然认为,地衣的起源只能追溯到第三纪(tertiary era)。当时,在莱茵兰褐煤和波罗的海琥珀中发现了第一批可确定的地衣遗骸,其中,在德国波罗的海琥珀中发现的绵腹衣(Anzia electra)(第三纪,下中新世,20 Ma)是已知最古老的地衣化石之一,已经进化出具有狭窄叶瓣的小型叶状体结构、海绵层(spongy layer)、双层髓质(medulla)和假根。
可以肯定的是,地衣并非一开始就长成了我们如今所看到的样子,而是经历了一个进化过程。对于该过程,我们目前所知的只是冰山一角。[21]
在地衣的组成成分中,蓝藻(cyanobacteria)*早在3800 Ma就已出现。作为已知最古老的陆地生物,它们的遗迹现存于澳大利亚等地发现的叠层石(stromatolites)上。而绿藻的起源则要追溯到真核细胞进化出叶绿体,这些遵循内共生关系(详见《共生和进化:真核细胞的起源》)的细胞器让植物得以进行光合作用。
作为地衣的另一种组成成分,真菌很难成为化石,目前已知首个真正意义上的真菌出现于前寒武纪(Precambrian)。发现于南非的前寒武纪(-2700到-2300 Ma)化石表明,真菌和藻类之间可能存在某种联系。自那之后,真菌便转向了与植物界并行不悖的发展之路,菌根(mycorrhizae)、地衣等共生体均代表了真菌与光合生物长期相辅相成的进化成果。
在中国贵州省独山县的化石地层中,发现了埃迪卡拉纪(Ediacaran period)早期(635-551 Ma)的地衣遗迹。这些保存极为完好的微化石呈现出与细胞相连的细丝,经分析,这些细胞与今天的蓝藻或绿藻十分相似。在英国发现了中奥陶世(Middle Ordovician)(460 Ma)的生物化石,疑似为某种地衣化真菌(Cosmochlaina),在电子显微镜下呈现出一个完整的皮层和次级皮层,但并未发现光合共生体。[22]
此外,在英国还分别发现了下泥盆纪时代(Lower Devonian)的沙罗普氏氯霉菌(Cyanolichenomycitis devonicus)和环核菌(Chlorolichenomycitis salopensis)化石,二者具有与现存地衣相似的结构,因此被认定为目前发现的年代最早的地衣。其中,前者表现为异质地衣体,表面覆盖有厚皮层,髓层中充满了蓝藻,类似于现在的念珠藻属生物(Nostocs)。这些地衣化石属于盘菌亚门(Pezizomycotina),其解剖结构与现在的茶渍纲地衣(Lecanoromycetes)相似[23]。
相比于目前公认的地衣样本,地衣共生的历史可能要悠久得多。由于地衣各组分难以形成化石,且发现的样品往往质量很差,相关研究因此阻力重重。然而,学界仍在持续改进现有的调查方法,如年代测定、分子生物学等,这些行动有望为未来地衣化石领域的发现提供有价值的信息。
在泥盆纪时期,各种微生物和地衣定殖于适宜气候区的岩石或土壤表面,这极有可能是生物征服陆地的第一步(详见《第一批陆地生态系统》)。现存的生物结皮让我们了解到地衣定殖的一些原始形式,但除此之外,还可能存在许多其他形式。目前,在最极端的生态系统(如北极、南极、高山、沙漠和草原)中,由于环境条件恶劣,地衣极少面临,或者说几乎不会面临来自高等植物的竞争压力,因而能够占据主导地位。微生物和地衣活动也在某种程度上改变了岩石和沉积物的状态,使得高等植物能够在此定居。随后,地衣还会参与土壤的形成,在其上扎根生长,并通过自身的有机碎屑实现自给自足。
5. 总结
除了深海、活体动物组织和严重污染地区,地衣可以在所有环境中生长。
地衣的共生比人们过去认为的要复杂得多:地衣体的结构不仅包括真菌+绿藻和(或)蓝藻两种基本伙伴关系,而且还涉及多种细菌,以及最近发现的单细胞真菌。
地衣可以复生:干燥后,一旦补充水分,它们就能恢复生机。
在只有藻类存在的情况下,真菌产生地衣分泌物,促使地衣固定在基质上,且能够起到保护作用。
地衣生长非常缓慢,有些地衣可以存活几个世纪。
地衣是开拓者,有能力在营养极度短缺的基质上定居,并能承受极端的温度和光照条件。
已知的第一批地衣可以追溯到泥盆纪。
参考资料和注释
封面图片:洛塔雷高山花园(Lautaret alpine garden)的岩石为丽石黄衣所覆盖(前景);拉梅耶山(the Meije)(海拔3984米)(背景)。[照片来源:© J. Joyard & M. Neuburger]
[1] 有时,真菌可以像腐生植物(saprophyte)一样从环境获取有机物,或像寄生虫一样生活在另一种地衣上。
[2] 多元醇在植物世界中广泛存在,真菌、地衣和许多藻类中都含有多元醇。地球上大约30%的初级碳来自植物和藻类合成的多元醇。在植物中,多元醇有助于生物体产生对非生物和生物胁迫的耐受力,并且是碳骨架的一种运输和储存形式。
[3] 有些地衣代谢物是有毒的。例如,杂髓地衣(Letharia vulpina)分布于高海拔地区,是一种具有皮层的地衣,外观呈现芥末黄色,主要生长于落叶松和石松上。这种地衣能够分泌有毒物质狐衣酸(vulpinic acid),因此曾被混在诱饵中用于消灭狼和狐狸。顺带一提,在诱饵中加入碎玻璃会增强猎杀的效果。
[4] 地衣自中世纪以来就作为定香剂而闻名,时至今日仍然广泛用于制香业。拟扁枝衣(Pseudevernia furfuracea)和常被误称为“橡苔”的栎扁枝衣(Evernia prunastri)是两种主要生长在中欧的地衣,因含有高浓度的黑茶渍素(atranorin),具有固香作用而遭到大量采收,并进口到法国格拉斯。当地香水厂会提取一种名为“橡苔净油”的浓缩物,用来调制多种香水。每年,法国南部和摩洛哥等国家和地区都会收获6000-8000吨地衣。如此密集的采收威胁着这类地衣的生存。理想的做法是用合成产品替代它们,但截至目前,这些地衣复杂的生化性质依然无人能够复刻。
[5] 自古以来,人们就从地衣中提取各种染料,呈红色的染料衣属地衣(Roccella)就是原料之一。其他地衣或像石脐属地衣(Umbilicaria pustulata)一样呈现棕红色、或如杂髓属地衣(Letharia vulpina)和皱梅属地衣(Flavoparmelia caperata)一般呈现黄色、或同黄衣属地衣一般(Xanthoria parietina),呈现橙黄色或粉红色、或像各类石蕊属地衣(Cladonia)一样呈现绿色。根据测试溶液的酸度或碱度而变成红色或蓝色的“向日葵”溶液(即石蕊试剂)是从染料衣属地衣、树形属地衣(Dendrographa)等多种地衣中提取的。尽管目前化学染料已经非常普及,但一些爱尔兰花呢仍坚持用地衣染色。
[6] Grube, M., Cernava, T. Soh , J., Fuchs, S., Aschenbrenner, I., Lassek, C., Wegner, U., Becher, D., Riedel, K., Sensen, C.W. & Berg, G. 2015. Exploring functional contexts of symbiotic sustain within lichen-associated bacteria by comparative omics. ISME J. , 9, 412–424.
[7] 与地衣有关的细菌极其丰富的多样性,以及其在地衣共生中的基本功能表明,在逻辑上,微生物群(即生活在生物体内或与生物共同生活的所有细菌)的概念可以很好地扩展到地衣。
[8] Spribille, V. Tuovinen, P. Resl, D. Vanderpool, H. Wolinski, M.-C. Aime, K. Schneider, E. Stabenheimer, M. Toome-Heller, G. Thor, H. Mayrhofer, H. Johannesson & McCutcheon, J.-P. 2016. Basidiomycete yeasts in the cortex of ascomycete macrolichens. Science, 353, (6298), 488-492.
[9] 与之相反的是,叶状地衣和枝状地衣的生长速度更快。目前,地衣的生长速度纪录为每年10厘米,是由一株树花属(Ramalina)地衣保持的。
[10] Beschel, R. 1957. Lichenometrie im Gletschervorfeld. Jahrbuch des Vereins zur Schutze des Alpenflora und Tiere. 22p.
[11] 粘液是一种植物分泌物,由多糖组成,与水接触后会膨胀成胶状粘稠物。
[12] Aubert, S., Juge, C., Boisson, A.-M., Gout, E. & Bligny, R. 2007. Metabolic processes sustaining the reviviscence of lichen Xanthoria elegans (Link) in high mountain environments. Planta, 226, 1287-1297.
[13] Bachereau, F. & Asta, J., 1997. Effects of solar ultraviolet radiation at high altitude on the physiology and the biochemistry of a terricolous lichen Cetraria islandica (L.) Ach. Symbiosis, 197-217; Bachereau, F. & Asta, J., 1998. Effects of solar ultraviolet radiation at high altitude on the phenolic compounds contents of Cetraria islandica (L.) Ach. Ecologie, 29(1-2), 267-270.
[14] Vera J.P., Horneck, G., Rettberg, P. & Ott, S. 2004. The potential of the lichen symbiosis to cope with the extreme conditions of outer space II : germination capacity of lichen ascospores in response to simulated space conditions. Adv. Space Res. 33, 8, 1236-1243.
[15] Scheidegger, C. (1995). Early development of transplanted isidioid soredia of Lobaria pulmonaria in an endangered population. Lichenologist, 27(5), 361-374.
[16] Galsomiès, L., Robett, M. & Orial, G. 1999. Interaction lichens-roche sur monument historique en granite. Bull. Inf. As. Fr. Lichenol. Mémoire n° 3, Grenoble, 35-42.
[17] Asta, J. & Letrouit, M.A. 1995. Observations on the early growth of Rhizocarpon geographicum thalli. Herzogia, 11, 229-252.
[18] Asta, A. et Souchier, B. 1999. Lichens et pédogenèse : dynamique de la végétation et études micromorphologiques de l’interface-lichen-sol. Bull. Inf. As. Fr. Lichénol. Mémoires n°3, 29-34.
[19] Asta, J., Orry, F., Toutain, F., Souchier, B. & Villemin, G. 2001. Micromorphological and ultastructural investigations of the lichen-soil interface. Soil Biol. Biochem. 33, 323-337.
[20] Scheidegger, C. 2015. Touche pas à ma croûte ! Année Internationale des sols. Fiche_croute_biologique.pdf
[21] Farou, J.L. 2017. Sur la piste des premiers lichens. Bull. Inf. As. Fr. Lichénol., 42, 2, 240-248.
[22] Edwards, D., Axe, L. & Honegger, R. 2013. Contributions to the diversity in cryptogamic covers in the mid-Paleozoic: Nemato thallus revisited. Bot. J. Linn. Soc. 173, 505-534.
[23] Honegger, R., Edwards, D. & Axe, L. 2013. The earliest records of internally stratified cyanobacterial and algal lichens from the Lower devonian of the Welsh Boerderland. New Phytol., 197, 264-275.
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: ASTA Juliette (2024年3月14日), 地衣,令人惊叹的开拓者, 环境百科全书,咨询于 2024年4月19日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/lichens-pioneering-organisms/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。

