图3. 烟草植株。当它的叶子受到掠食者攻击时,烟草会像许多植物一样合成对食草动物有毒的酚类化合物来保护自己。此外,一如桤木、苦艾或糖枫,它会释放一种警报信息素(挥发性化合物),诱导附近同种植物合成相同的信息素。[来源:福雷斯特(Forest)和金·斯塔尔(Kim Starr),CC BY 3.0,通过维基共享资源]
[4] Teyssèdre A., 2006. Les clés de la Communication animal. Delachaux et Niestlé, Paris.
[5] MacArthur R.H. & E.O. Wilson, 1967. The theory of island biogeography. Princeton Univ. Press, Princeton, NJ.
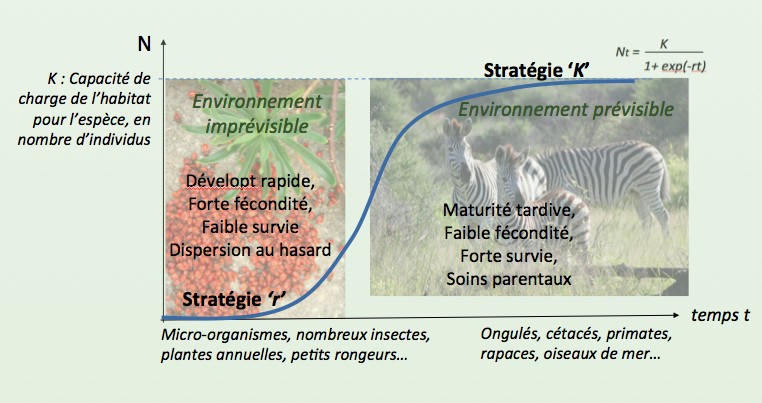
[6] Pianka E.R., 1970. On r and K selection. Am. Nat. 104:592-597.
[7] Reznick D., M.J. Bryant & F. Bashey , 2002. R- and K-section revisited: The role of population regulation in life history evolution. Ecology 83(6):1509-1520.
[8] Beaumont H.J.E., Gallie J. et al. 2009. Experimental evolution of bet hedging. Nature 462:90-92.
[9] Botero C.A., F.J. Weissing, J. Wright & D.R. Rubenstein, 2015. Evolutionary tipping points in the capacity to adapt to environmental change. Proc. Natl. Acad. Sci. USA 112:184-189.
[10] Starrfelt and Kokko 2012. Bet hedging: a triple trade-off between means, variances and correlations. Biol. Rev. 87, pp. 742-755.
[13] Teyssèdre A., 2018. Les mammifères face aux changements globaux. Regards et débats sur la biodiversité, SFE, regard R80b, June 2018.
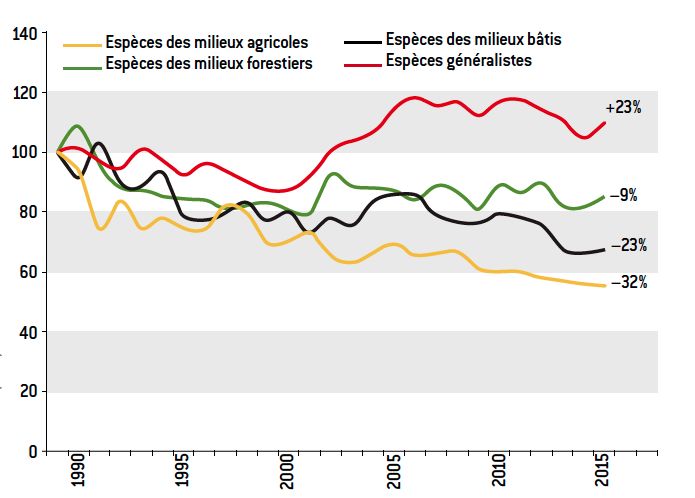
[14] McKinney M.L. & J.L. Lockwood, 1999. Biotic homogenization: a few winners replacing many losers in the next mass extinction. T.R.E.E . 14:450-453
[15] Clavel J., Julliard R. and V. Devictor, 2010. Worldwide decline of specialist species: toward a global functional homogenization? Front. Ecol. Environ. 2:222-228.
[16] Thomas C.D., 2015. Rapid acceleration of plant speciation during the Anthropocene. Trends Ecol. Evol. 30:448-455.
[17] Julliard R., J. Clavel et al. 2006. Spatial segregation of bird specialists and generalists in bird communities. Ecol. Letters 9:1237-1244.
[18] Clavel J., 2011. L’Homogénéisation biotique. Regards et débats sur la biodiversité, SFE, regard n°16, April 2011.
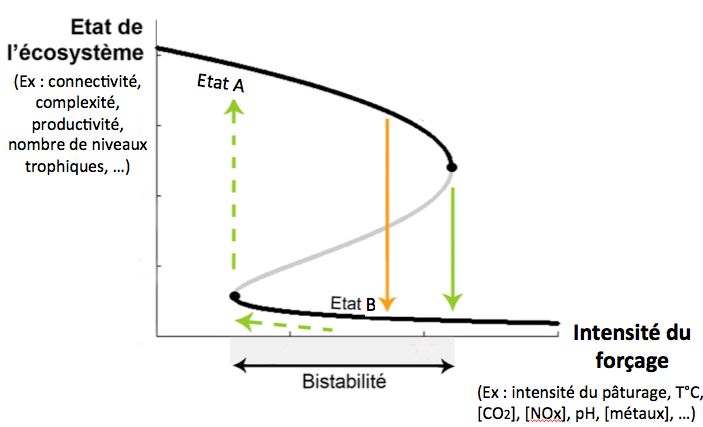
[19] Folke C., S. Carpenter et al. 2004. Regime shifts, resilience and biodiversity in ecosystem management. Ann. Rev. Ecol. Syst. 35:557-581.
[20] Cardinale B. et al. 2012. Biodiversity loss and its impact on humanity. Nature 486:59-67.
[21] Haberl H., 2007. Quantifying and mapping the human appropriationof net primary production in earth’s terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 104:12944-12947.
[22] Gaston K.J., T.M. Blackburn, K. Klein Goldewijk, 2003. Habitat conversion and global avian biodiversity loss, Proc. R. Soc. Lond. B 270:1293–1300.
[23] Smil V., 2011. Harvesting the Biosphere: the Human Impact. Pop. Dev. Rev. 37(4):613-636.
[24] Teyssèdre A. & D. Couvet, 2007. Expected impact of agriculture expansion on the world avifauna. C. R. Acad. Sci. Biol. 330:247-254.
[25] Pauly D, V. Christensen V, J. Dalsgaard J, R. Froese & F.S.B. Torres, 1998. Fishing down marine food webs. Science 279:860-863.
[26] Watson R. & D. Pauly, 2001. Systematic distortions in world fisheries catch trends. Nature 414(6863):534-536.
[27] Folke C., S. Carpenter et al. 2004. Regime shifts, resilience and biodiversity in ecosystem management. Ann. Rev. Ecol. Syst. 35:557-581.
[28] Cardinale B. et al. 2012. Biodiversity loss and its impact on humanity. Nature 486:59-67.