




色素色彩的起源
PDF1. 从纳观尺度的电子与光子解析蝴蝶色彩多样性

所有分子都与电磁辐射(即光,范围从高能伽马射线、X射线到低能无线电波)相互作用。但并非所有分子都显色,因为人眼仅能感知可见光波长(400至700纳米)。在生物界,有机分子主要吸收远紫外线波长(100至300纳米),绝大多数在我们看来是透明的;这一普遍特性也解释了为何生物体分子易被这些能量较高的波长降解(就DNA而言可能导致突变)。
然而,色彩在我们所感知的世界中无处不在(如图1所绘蝴蝶的绚丽色彩所示),这引出一个问题:为何某些分子能吸收比平均波长更长(因而能量更低)的光?
这些具有特殊吸收特性的分子被称为色素[1]。要理解色素的独特性,需解释物质与波长200至1000纳米(从紫外线到近红外,含可见光)的光相互作用的原理。
2. 光子与电子的相互作用
光由称为光子的粒子组成,光子携带特定能量(能量与波长成反比)。物质由分子构成,分子由原子相互作用形成,原子则由原子核和电子组成。正是这些带负电的粒子,能与目标波长范围内的光子相互作用。
电子在静止时具有自身能量,且仅当吸收的能量精确等于其初态与终态能量差时,才能跃迁至少数更高能级。显然,并非所有光子都能激发所有电子:电子仅当光子提供恰好足够的能量跃迁至高能态时,才能吸收该光子。
类比而言,电子如同无线电天线,其特性使其仅能接收特定频率,因为只有极小部分频率会与天线相互作用。以DNA分子为例,其电子仅能被紫外线波段光子对应的能量激发。光谱中其他光子(尤其是可见光波段)会直接穿透DNA分子,不发生任何作用。
因此,分子要显色,其电子必须处于高于平均的起始能级,或能跃迁至低于平均的终态能级,因为可见光光子能量低于紫外线光子。然而,电子本质相同,孤立来看色素中的电子与DNA中的并无二致。实际上,是化学环境改变了它们的能级。
3. 色素:与众不同的分子
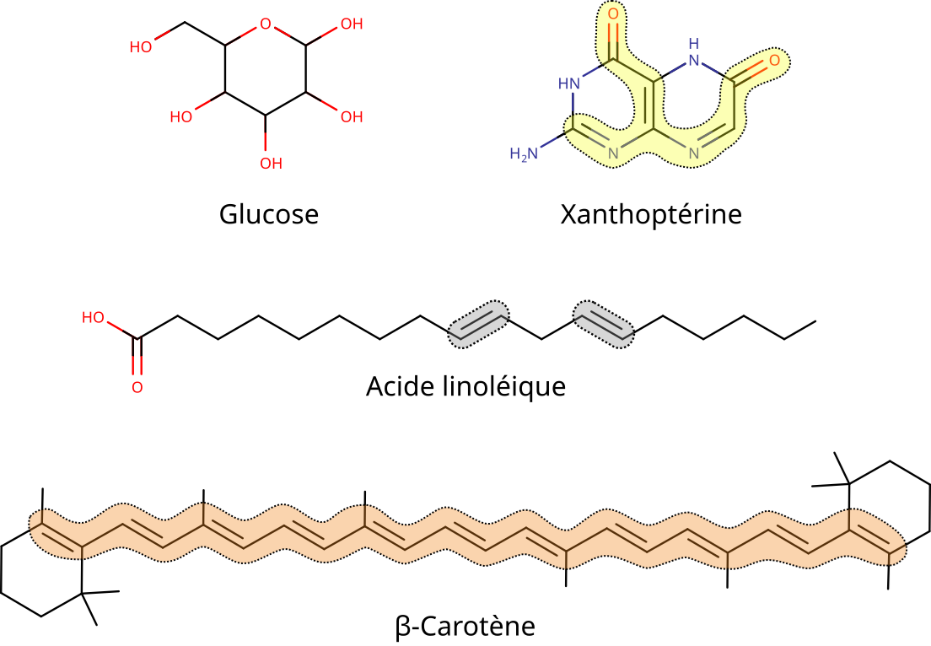
比较色素与透明有机分子的结构(图2)可知,原子间的特定构型是改变电子性质的关键(需注意以下参数并非充分必要条件,亦不具排他性):
- 足够长度的单双键交替,即共轭双键;
- 含有共轭键的芳香环结构;
- 直接连接共轭键/芳香环的杂原子(氮、氧、硫等)的存在与位置;
- 与蛋白质、金属等其他化合物的相互作用。
4. 色素色彩多样性的电子起源
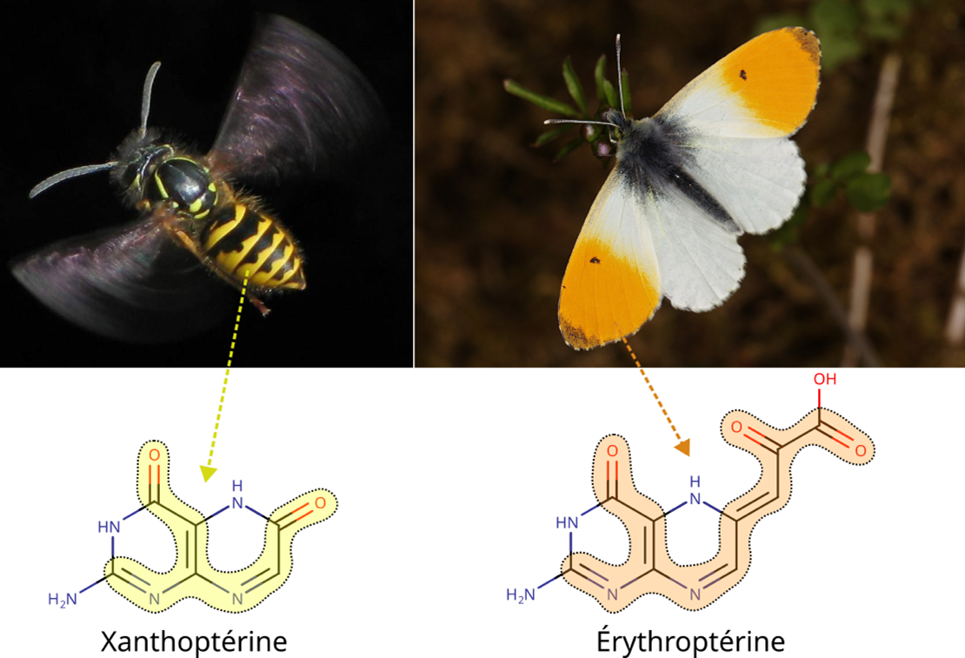
使色素显色的特性也决定了其不同色调。实际上,上述任一参数的变化都可能导致色素电子吸收不同波长的光。我们感知的色素颜色是其吸收色的补色,这意味着吸收紫光的黄蝶呤(黄蜂黄条纹)与吸收蓝光的红蝶呤(奥罗拉蝶橙斑;图3)呈现不同色彩。
我们可进一步发现,此处将波长范围限定在400-700纳米,仅因人类眼睛对此范围之外的光不敏感(人类视网膜中的色素也仅对特定波长范围响应,这同样取决于其电子特性——形成闭环!)。然而,其他生物未必受此限制。例如,蜜蜂和鸟类常能感知近紫外线(350-400纳米),某些蛇类则能看见近红外线(700-1000纳米)。
因此,“可见”与“不可见”并无明确分界,仅是有机分子特性决定的光谱吸收梯度(就此而言,色素除愉悦人眼外,与其他有机分子并无本质不同)。
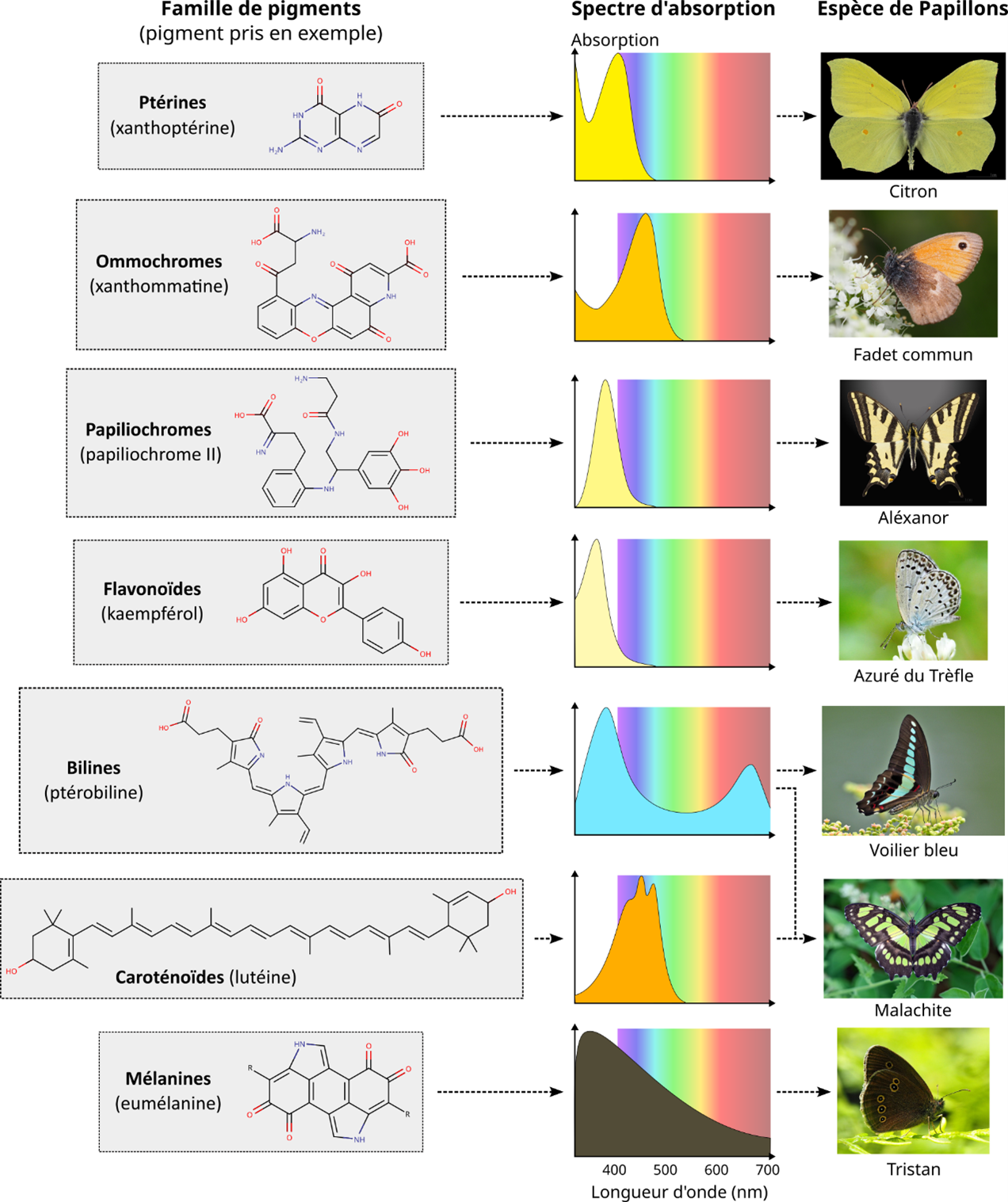
实际上,通过紫外-可见-红外光谱技术获取色素的吸收光谱(图4)可见:色素并非吸收单一波长,而常吸收光谱中多个点或连续区间,围绕均值形成宽窄不一的吸收峰(图4)。这是因为色素电子受其复杂环境(溶剂、基质、蛋白质等)影响,能吸收一系列相邻波长。
最终,色素电子的能级决定其在紫外-可见-红外波段的整体吸收光谱,结合本文未涉及的其他着色机制,共同赋予生物体可见色彩(图4)。
拓展阅读
- Figon, F. & Lorin, T. (2019) La coloration des Animaux : éléments de physique et de chimie. Planet-Vie. https://planet-vie.ens.fr/thematiques/cellules-et-molecules/biophysique/la-coloration-des-animaux-elements-de-physique-et-de (in French)
- Valeur, B. & Bardez., E. (2015) La lumière et la vie, une subtile alchimie. Paris : Belin. (In French)
参考资料及说明
题图:君主斑蝶(Danaus plexippus)[图片来源:图卢兹自然历史博物馆,CC BY-SA 4.0许可,经由维基共享资源]
[1] 从生物化学角度看,色素可直接来源于食物(例如源自植物叶绿素的绿色胆色素、完整保留的植物类黄酮或由生物体自身代谢产生(例如黑色素、蝶呤和眼色素/凤蝶色素分别由酪氨酸、鸟嘌呤和色氨酸等氨基酸或核酸碱基合成)。