森林与全球环境变化

数千年来的大规模林地清理和集约采伐已显著减少了法国的森林面积。尽管自19世纪起就开始实施重新造林措施,人类活动仍然对树木和森林造成了巨大压力。自工业革命以来,空气污染和大气中温室气体浓度上升引发了一场不断加剧的气候变化。有毒污染物、强烈或反复的干旱、高温以及风暴,都给树木带来了严重影响。这些影响导致树木生长显著减缓、叶片明显脱落、枝条稀疏以及死亡率上升。树木天然防御能力的削弱使其易受昆虫、真菌或其他病原体等生物侵害。这些侵害进一步加剧了树木的衰退,对森林林分的健康和恢复力造成了长期损害。
1.人类活动对森林的影响
地球上的人类活动导致了(i)大规模的环境污染(空气、水体、土壤),(ii)主要由大气中二氧化碳浓度持续上升引起的气候变化 [1],以及(iii)对生物多样性的严重破坏。
因此,自工业革命开始以来,大气中的二氧化碳含量已增加50%,从1800年的280 ppm上升至2024年的424 ppm。这尤其导致了1850年至2024年间全球平均地表温度不可避免的上升,幅度约为1.5°C至1.6°C,从而加剧了极端事件(干旱、热浪等),影响了森林、植物、动物和人类的健康在内的所有生态系统。
然而,早在1800年,亚历山大·冯·洪堡(Alexander von Humboldt)以及随后19世纪的其他生态学先驱就曾警告森林砍伐对气候的影响,并倡导人类、自然、树木与森林之间建立和谐关系 (可参见焦点 生态学先驱 )。
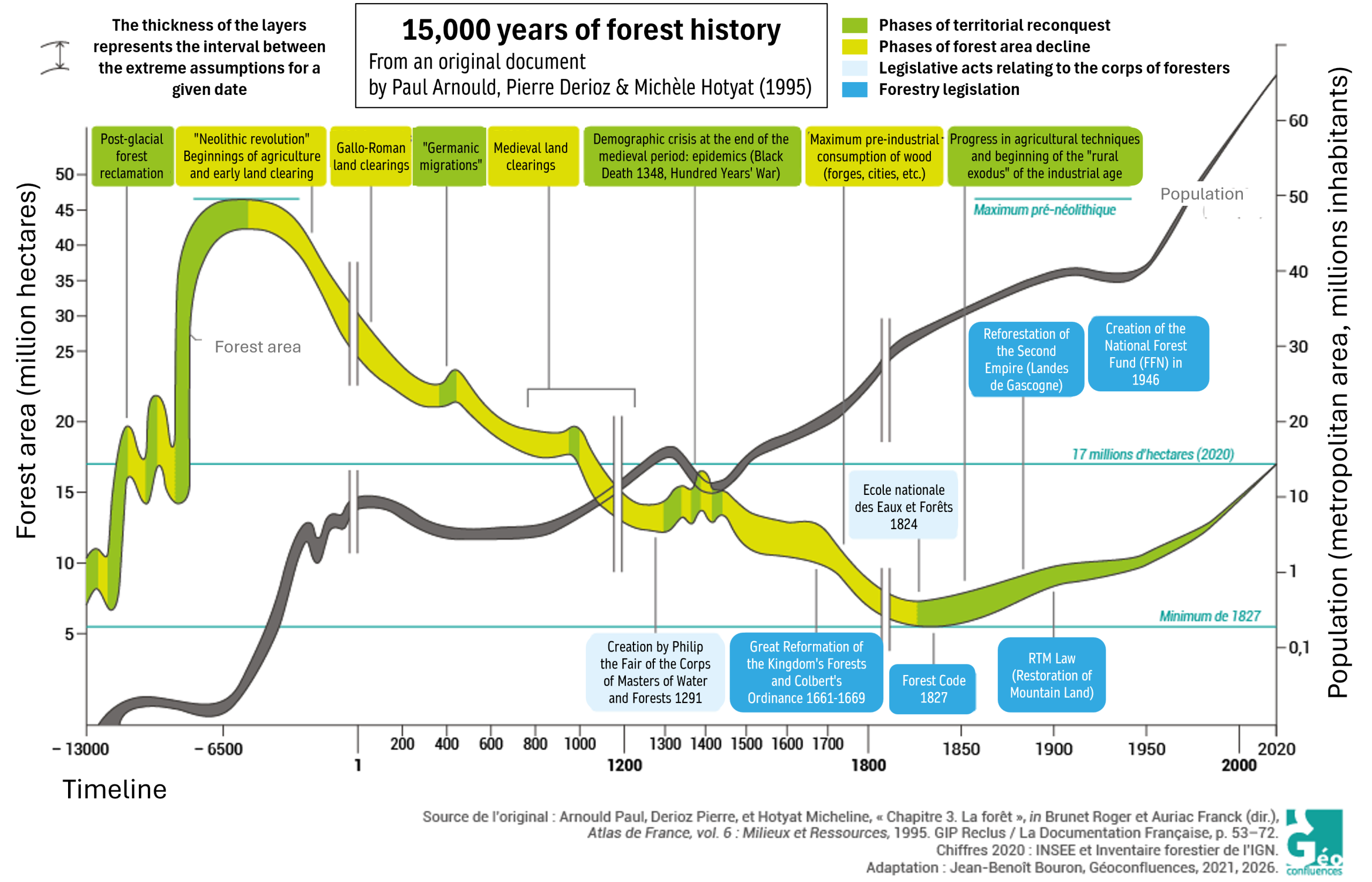
自欧洲末次冰期结束后,得益于更为适宜的气候,森林覆盖面积显著增加(图1)。随后,从农业出现(约公元前6000年)开始,尽管有过一些短暂的恢复期(罗马帝国灭亡、百年战争、黑死病——图1),法国的森林覆盖率仍因大规模砍伐和刀耕火种的做法而持续下降,直至19世纪[2]。科尔贝于1669年颁布的《森林法令》仅暂时稳定了局势,但随后的主动植树造林和保育努力使森林覆盖率显著上升,直至20世纪中期。
第二次世界大战后,国家森林基金开展的重新造林工作,以及农业与牧业用地的大规模弃置(这些土地自然演替为林地),共同加速了森林的恢复,这一进程一直持续到21世纪,从未中断。
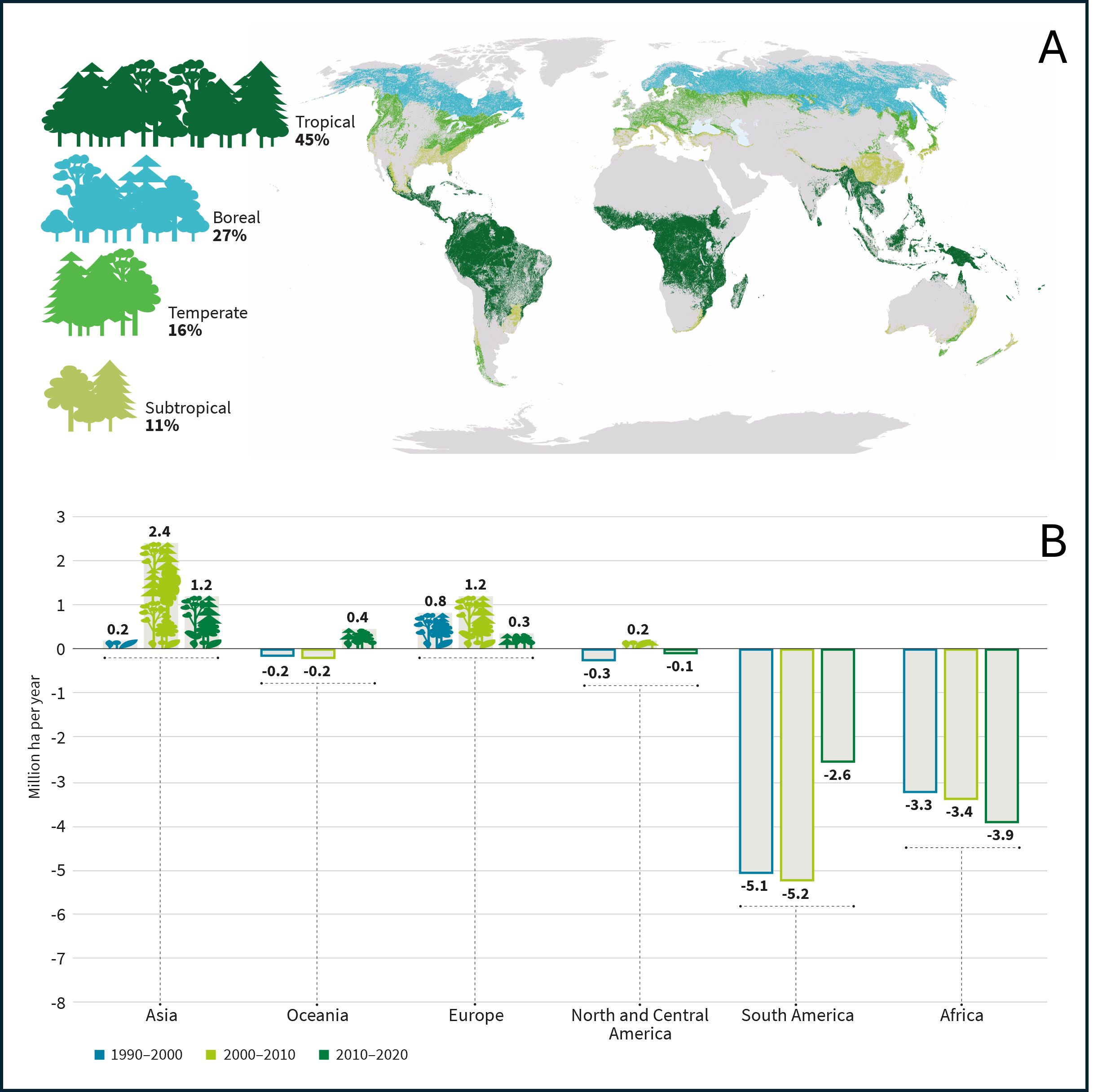
全球范围内,森林可分为四个气候区(图2)[3];其中面积最大的是热带森林,占全球森林总面积近一半(图2A),该区域的特点是毁林现象十分严重:
- 过去三十年间,南美洲和非洲的热带森林面积出现了急剧下降(图2B),主要原因是森林火灾以及巴西亚马逊地区和刚果民主共和国为农业目的(自给性农业或工业化农业,如大豆种植等)及木材采伐所导致的毁林行为。
- 在亚洲,印度尼西亚正通过用油棕种植园取代森林,积极推动这一面积减少趋势。
这些蓄意毁林行为占全球向大气中排放的二氧化碳总量的58%。
在欧洲,与积极的再造林政策相关的管理改善使得1990年至2010年间森林覆盖率显著增加,碳储量也随之增加,但这一增长在2010年至2020年期间有所放缓(仅增加30万公顷,而此前十年增加了120万公顷,图2B)。
2.目前导致森林衰退的因素
干旱、热浪、风暴[4]和病害的共同作用,抑制了树木生长并增加了树木死亡率。自19世纪以来,各大陆均观测到与严重干旱相关的森林枯梢现象,而近几十年来,这一现象变得更加频繁和广泛[5][6]。
森林火灾(无无论是人为纵火还是由诱发火灾发生的气候条件所引起)进一步加剧了这一效应,对全球各地造成毁灭性破坏,包括:(i)热带地区(亚马逊等);(ii)北美洲(美国、加拿大);(iii)欧洲(法国、意大利、西班牙、葡萄牙、希腊)[7];(iv)北方地区(俄罗斯等);(v)以及澳大利亚和印度尼西亚。
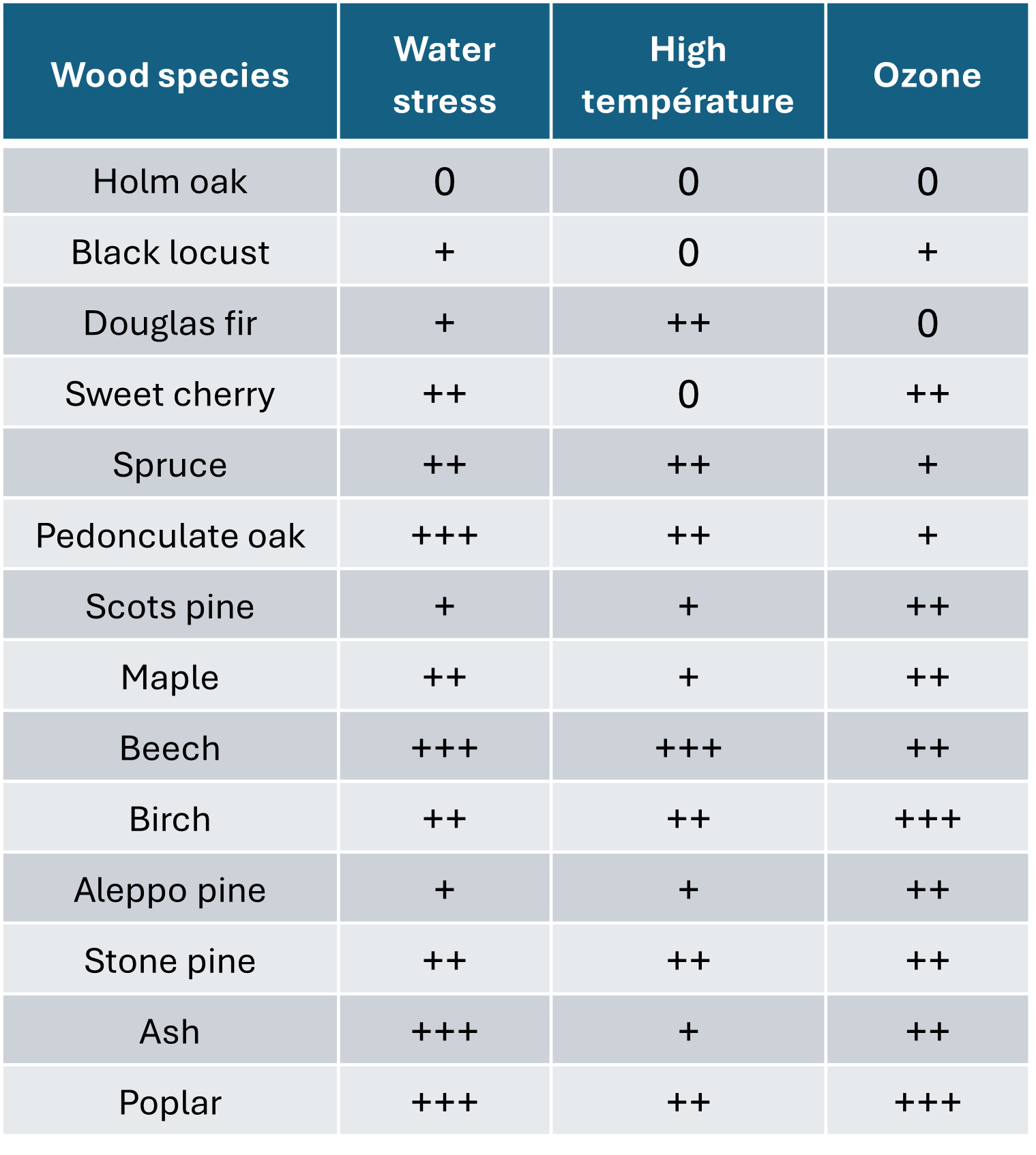
除了上述所有现象之外,还存在大气污染问题。欧洲最常见的森林树种对各种胁迫表现出不同程度的敏感性(表1)。此外,我们还必须考虑生物胁迫(真菌、昆虫、细菌),这些胁迫对已被气候变化削弱的树木更具破坏性。
表1. 不同森林树种对各种胁迫的敏感性。 改编自 Claessens H, 2016[8]。0:耐受;+:不敏感;++:敏感;+++:非常敏感。
这种衰退的后果是,作为主要碳汇的森林吸收二氧化碳的能力正在下降。1990年至2010年间,森林的碳储存能力下降了约30%[9]。在亚马逊地区,这一下降被认为发生在短短20年间(2000年至2010年)[9]。森林火灾通过释放大量二氧化碳[10],削弱了森林碳汇,从而形成了一个恶性循环。
在法国,根据最新的森林清查数据,年碳固存量已从2005-2013年间的6300万吨二氧化碳下降至2014-2022年间的3900万吨[9]。
大规模的热带森林砍伐,加上日益加剧的气候影响(干旱、火灾、树木死亡),正在严重削弱森林在全球范围内作为碳汇的功能,包括在欧洲和法国,这些地区的森林恢复力正在下降。
3.树木与污染

自人类过上定居生活之日起,以取暖和开垦为目的的用火行为就造成了空气污染,而自18世纪中叶起的工业革命则使这一问题大大加剧(可参见焦点 空气污染与树木)。
直至20世纪80年代,空气污染主要以工厂和家庭燃煤产生的二氧化硫为主。随后,臭氧成为主要污染物 (可参见 空气污染 )。这种次生污染物是在紫外线作用下,由氮氧化物、碳氢化合物残留物以及源自道路交通(以及某些植物,特别是在高温干旱期)的挥发性有机化合物形成的。研究已证明,这些污染物对所有植物均产生有害影响,会损害植物叶片并导致生物量损失[11],[12],[13] (可参见 空气污染对植被有什么影响 )。
当东风将莱茵平原的污染空气吹向孚日山脉时,臭氧对树木的可见影响便显现出来。云杉的针叶出现明显的黄化(图3A)。在希腊,阿勒颇松的针叶上呈现出典型的臭氧损伤斑点,称为“褪绿斑驳”(图3B)。

在欧洲和北美,大气臭氧浓度正趋于稳定,而在亚洲则持续上升。在法国,臭氧仍是每年夏季的主要污染物(图4)。根据欧洲环境署的最新报告,2022年,32个成员国的森林总面积中有62%暴露于超过临界阈值的臭氧浓度之下。
森林的脆弱性源于臭氧与气候变化影响之间的相互作用[14]。一项近期研究表明,在亚马逊地区,与干旱相关的火灾加剧了臭氧造成的损害,而臭氧的形成又因树木燃烧释放的氮氧化物而得到促进[10]。其结果是,大气中的二氧化碳增多既来自火灾的释放,也来自臭氧对光合作用的抑制。
4.干旱与热浪下的森林
4.1树木对干旱和高温的生理响应
全球变暖(i)以每十年数天的速度延长了生长季,表现为(ii)发芽提前和(iii)衰老延迟。这增加了树木的耗水量,也加剧了水分胁迫的风险。干旱和热浪事件也变得更加频繁。
水凭借水势梯度从根部输送到叶片(参见 植物对水孜孜不倦的追求 )。一棵胸径为45厘米的橡树每天可吸收约一百升水。当土壤变干时,这一水流可能中断,导致输导组织中发生空穴化(形成气泡)。在此之前,调节叶片蒸腾作用的气孔会关闭,以防止水分过度散失。但这样做会减少二氧化碳的进入,而二氧化碳是光合作用中生化反应的燃料(参见 光合作用中碳的路径 ),从而减缓树木的生长。
温度在光合作用中起着关键作用,每种森林类型都有其最适温度(参见 温度对光合作用的影响 ):
- 北方森林:最适温度约为 20 °C;
- 温带森林:最适温度约为 25–30 °C;
- 热带森林:最适温度约为 30–35 °C(林冠层通常接近 30 °C)。
当温度超过这些最适水平时,(i)光合酶会受到损害,(ii)叶绿素色素发生降解,导致叶片或针叶变色、继而干枯并脱落。
最后,由于高温和干旱这两种现象常常同时发生,要区分它们各自的影响已变得越来越困难。
4.2干旱与高温对森林的当前影响
在法国及其邻国,数十年来森林一直受到干旱的影响[15][16]。历史上曾发生过显著的干旱事件(如 1892–1897 年、1910–1917 年、1922–1927 年、1946–1949 年、1955–1961 年、1976 年、1989–1991 年)。自 20 世纪末以来,这些事件的频率不断增加,并常常伴随强烈热浪同时出现。

2003年的夏季导致法国东北部橡树林年生长量下降25%。2003年夏季导致法国东北部橡树林的年生长量下降了25%。自那时起,与气候相关的胁迫在欧洲和法国都变得越来越频繁。在欧洲,尤其是2018年和2022年;在法国,2011年、2022年和2024年均出现了创纪录的年均或夏季气温,并伴有长期干旱。全球气温上升正增加世界各地树木受损的风险:在澳大利亚,过去几十年所有森林生物群落中的树木死亡率均显著上升[17]:
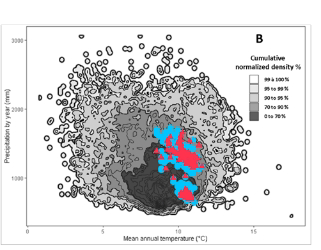
- 在法国地中海地区,2003–2007年的平均降水量亏缺(五年累计减少48%)仍是过去160年来的最高纪录。
- 在普罗旺斯,过去十年(2015–2024年)中有八年既缺水又非常炎热,加剧了自1990年以来持续上升的落叶程度:2024年,落叶树达52%,针叶树达42%(图5)。
在法国,落叶率和树木死亡率(图6)自2003年以来持续上升,降低了森林生产力,尤其是在树种分布区的南缘或山地物种的低海拔地区[18]。在异常年份,即使在树种分布区的核心地带也观察到了这一现象。例如,2019年法国东北部在经历了一月至九月降水量亏缺、同时伴有创纪录的平均气温(有记录以来第三最热年份)、两次破历史纪录的热浪以及异常日照(偏多10%–20%)之后,便出现了上述情况。
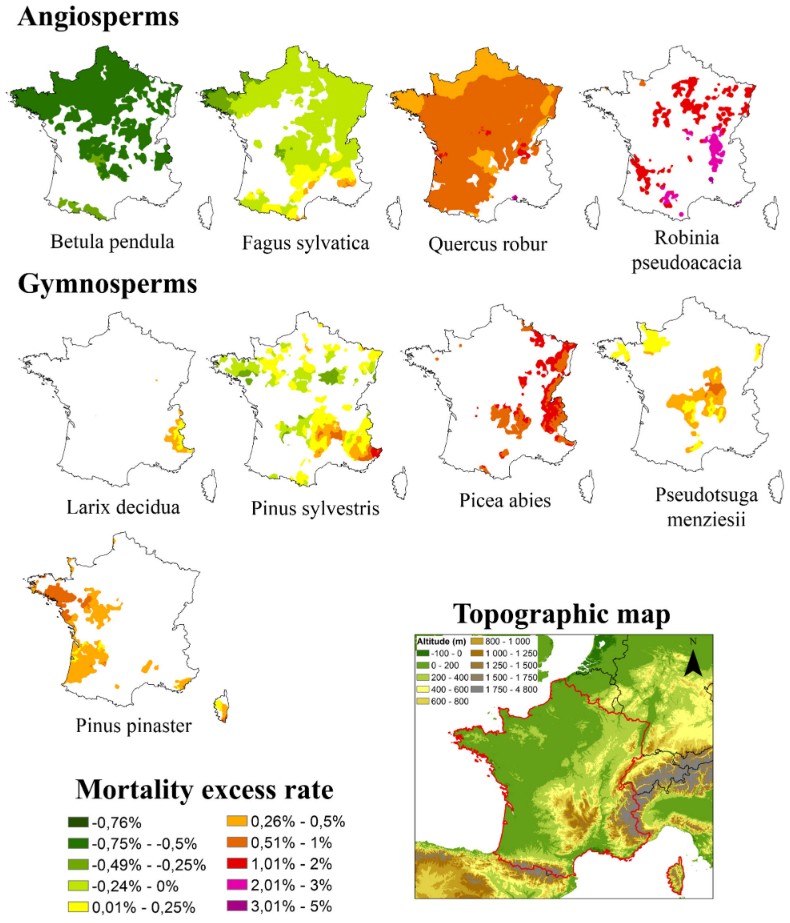
这些不利因素的共同作用导致土壤严重干涸,几乎所有树种都受到胁迫,使得本已很高的死亡率进一步加剧。一段时间以来,这些地区的森林排放的碳已经超过了其吸收的碳。就整个法国而言,过去十年间森林碳储量下降了40%。

面对干旱,森林生产力也因树种而异:
- 对于法国地中海地区的两种优势松树,就樟子松和阿勒颇松 [19][20]而言,其生产力在21世纪之交出现了显著下降(图7):
- 阿勒颇松(最耐旱的本土树种)在20世纪最初受益于气候变暖,随后出现了急剧逆转;2019年,其在普罗旺斯的平均落叶率达到了40%(气候胁迫加上机会性病原体)。
- 樟子松处于其分布区的下限,一个多世纪以来一直在快速衰退。
- 毛栎和冬青栎在20世纪的生产力增长了40%以上,但自2003–2007年以来,它们开始大规模死亡:
- 这些栎树中有三分之二的落叶率超过50%;
- 健康毛栎分布的下限已向高海拔地区推移了数百米,并向北推移了约100公里。
矛盾的是,一些被认为耐热或耐旱的树种(如阿勒颇松、冬青栎以及地中海灌木林中的优势灌木)正在遭受严重的枯梢病,尽管气候尚未超出它们已知的生物学极限。对此有多种解释:包括林分年龄、遗传因素、土壤退化导致肥力下降、生态系统系统性紊乱以及病原体引发的剧变。
多年反复出现的胁迫使森林无法恢复,从而降低了其恢复力。普罗旺斯栓皮栎林在反复遭遇火灾和干旱后出现的高死亡率和健康状况恶化就是一个很好的例子。这些森林在火灾和2003年热浪(此前几年为多雨年份)之后曾一度恢复,但从2005–2007年起开始遭受严重的枯梢病,而在过去50年中经历过多次火灾的地块,这种情况更为严重。2007年发生火灾时已是连续第五个干旱年,即使是在此前50年未发生过火灾的地区,林分也遭到了毁灭性打击。到2025年,由于过去十年间反复干旱,这一森林生态系统仍未恢复,死亡率居高不下,整体健康状况严重恶化[21]。
这种恶性循环影响着整个生态系统(植物、土壤、动物、微生物):反复干旱使生态系统对火灾更加敏感,而反复火灾又使生态系统对干旱和热浪更加脆弱。在法国国家层面,法国国家农业、食品与环境研究院(INRAE)的预测[22]表明:
- 从2030年起,火灾将增加(根据指标不同,增幅为13%至22%,指标包括过火面积、失控火灾数量、重大火灾数量)。
- 大型野火(>100公顷)的数量可能增加。在最坏的情景下,每年发生次数将从目前的约7次增至2050年的10次,到2090年可能多达20次。
从全球范围来看,预计到本世纪末,大型野火的发生次数将增加四倍。干旱和热浪事件的频率增加和持续时间延长将进一步加剧这一趋势,从而对森林生态系统构成长期威胁。
5.不同胁迫之间的相互作用
森林树种的衰退很少由单一环境因素引起。这是一个多因素作用的过程,通常是多种非生物和生物因素同时或接连影响的结果。这些因素之间复杂的相互作用最终导致了树种的衰退[23]。
一棵健康的树木能够产生防御分子,有效应对其通常面临的各种侵害。然而,当树木遭受主要非生物胁迫(如干旱、污染等)时,其防御能力会减弱,从而容易受到病原体或害虫的二次侵扰。反过来,一棵已经被寄生虫或疾病削弱的树木,也会对非生物胁迫更加敏感(图8)[16]。此外,非生物胁迫本身之间也会相互作用,例如火灾与干旱,或者干旱与热浪。
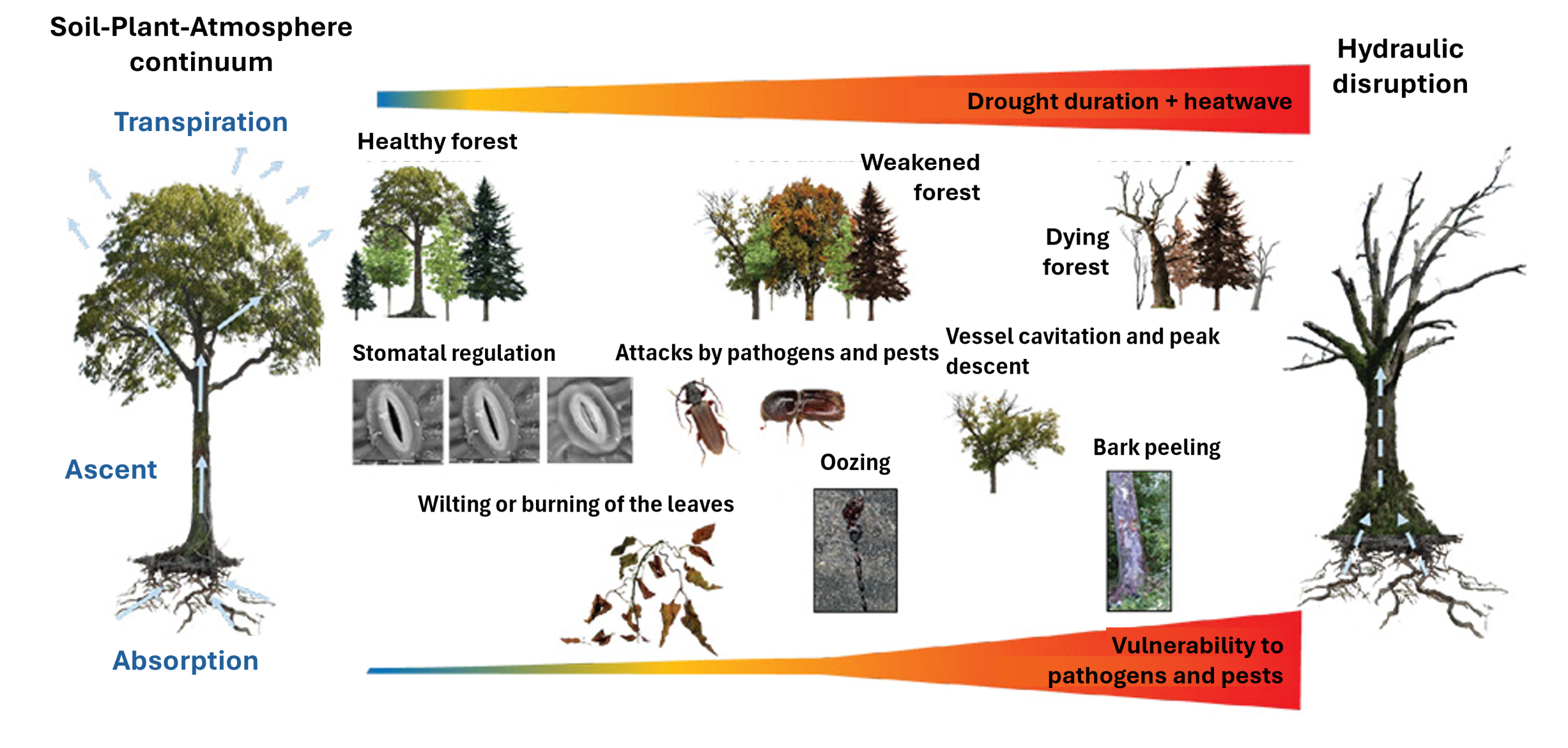
5.1非生物胁迫引起的萎蔫

北美地区暴露于非生物胁迫下针叶树的死亡情况。在 1950 至 1970 年代,洛杉矶东部山区的黄松因长期暴露于高浓度臭氧污染,遭受了松树小蠹的大规模侵扰,导致大量树木死亡(图9A) [24]。同样,在 1990 年代,北卡罗来纳州的米切尔山上,气候胁迫(严冬)、非生物胁迫(酸雨和臭氧)以及昆虫侵袭的共同作用,导致了针叶树的高死亡率(图 9B)。

地中海地区阿勒颇松近期的死亡情况。反复发生的干旱加剧了阿勒颇松的死亡,并且其死亡与地中海森林中的高臭氧浓度相关 [25].。2015年至2017年间,树皮甲虫(small beetles, 参见 4.2 和 焦点 生物性有害生物与森林衰退 )的侵袭加速了这一衰退过程。
中欧和东欧的云杉枯梢病。在过去约十年中,因干旱和热浪而衰弱的云杉树遭受了树皮甲虫的大规模侵袭,导致死亡率很高(图10),尤其是在法国东部和中欧地区。预计2018年至2024年间,欧洲的死木体积将达到约2.5亿立方米,超过1999年风暴造成的损失(欧洲为2亿立方米,其中法国为1.4亿立方米)。
影响法国和欧洲山毛榉及其他树种复杂因素。山毛榉及其他树种的衰退是多种主要非生物因素(气候、土壤、地形、海拔、坡向)和生物因素共同作用的结果,无论是在其最适分布范围的核心区域还是北部区域,情况均是如此(图11) [26]。这些树木原本适应曾经有利的气候,但它们在遗传、形态或生理上并未对近期的水分和热胁迫做好准备。它们容易受到土壤共生关系(菌根、细菌)紊乱、新病原体的入侵以及过去仅为隐蔽性地方物种的害虫流行性增殖的影响。
更广泛的生态系统影响。主要胁迫会扰乱整个森林生态系统,包括捕食者与猎物的平衡、营养级相互作用以及共生关系(尤其是土壤中的共生关系)。这会导致土壤肥力和生物多样性下降,从而使树木更加脆弱。从长远来看,由于不利的气候条件,种子传播者和传粉者等关键物种的减少会阻碍枯死树木的更新,并加剧森林衰退。
5.2生物胁迫的加剧
全球变暖极大地促进了生物害虫的繁衍,这些害虫成为森林衰退的主导因素。
- 蛀干昆虫和食叶昆虫
树皮甲虫是一种小型蛀干甲虫,它们在树皮下钻洞取食,在森林衰退中扮演着重要角色。全球变暖使它们每年能够多完成多达两个繁殖周期。这导致它们的种群数量呈指数级增长[27],并将其侵害活动延长至数月之久。由此造成的树木死亡为这些害虫的进一步增殖提供了更多机会,从而形成一个恶性循环。
另一个例子是松异舟蛾。这种害虫与法国东南部樟子松的衰退有关[28],冬季寒冷天气的显著减少,使该蛾虫既获得了很高的存活率,又延长了活动时间。五十年来,其种群密度不断增加,分布范围向高纬度扩展了300公里,向高海拔扩展了300米。
- 种子捕食者与繁殖受阻
山毛榉(以及某些栎树)的情况则具有矛盾性。这些树种以往的繁殖策略是将种子生产集中在间隔多年的年份中。全球变暖使其结果实变得更为规律,从而维持了以其种子为食的捕食者的高种群数量。这种加剧的捕食行为反过来又危害了它们的繁殖[29]。
- 生长季延长助长了真菌病害
在温带和寒冷气候区,病原真菌如今受益于显著延长的增殖期(早春和晚秋)。阿勒颇松的枝枯病在20世纪还只是一种低水平的地方性病害,如今已演变为流行病。这种病害会毁坏该树种的嫩枝,严重限制其生长。
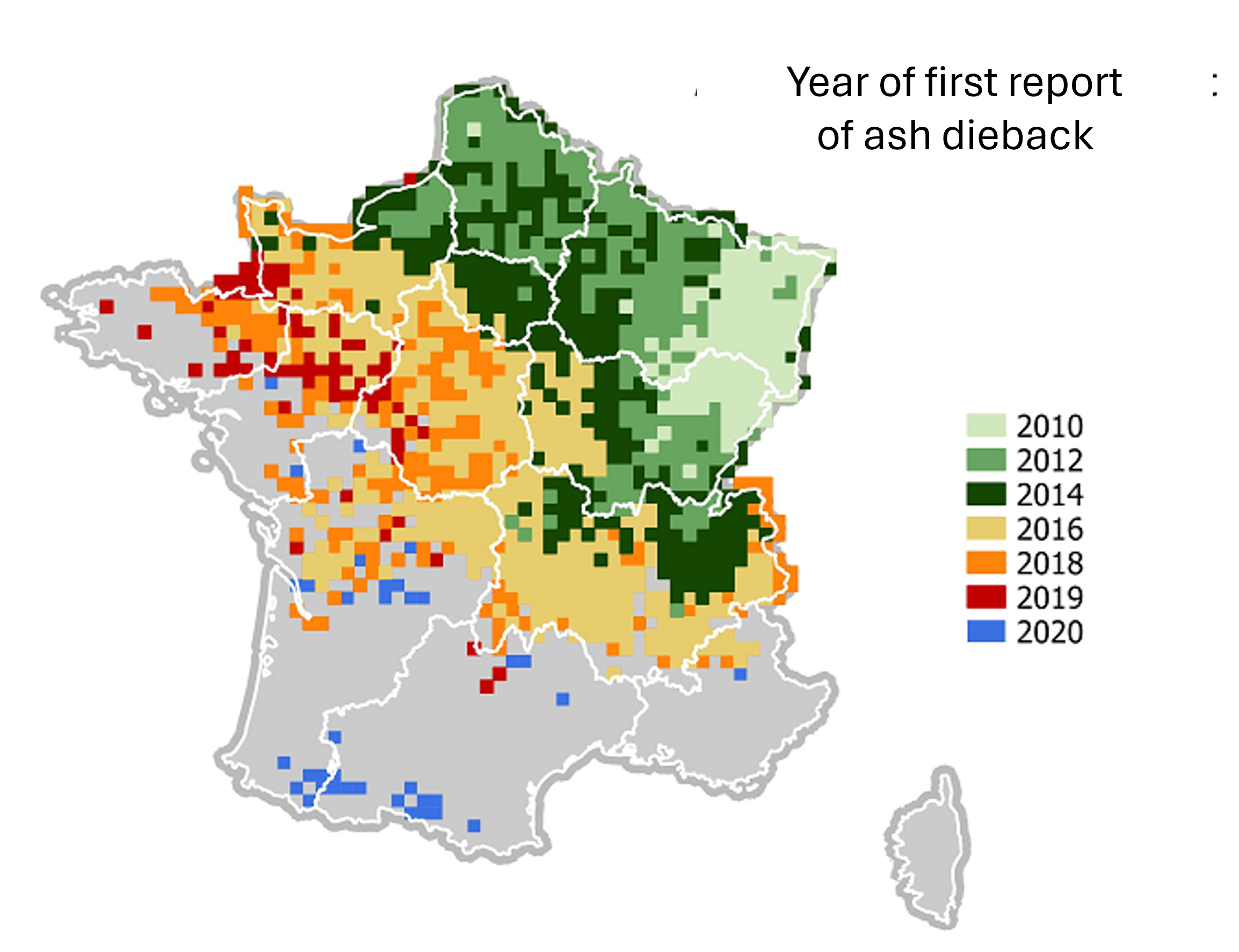
- 入侵性病原体与害虫(全球化因素)
随着全球化的发展,疾病和害虫的意外引入正在造成大范围的破坏。白蜡树枯梢病就是一个典型例子。这种疾病由真菌 Chalara fraxinea 引起。它从亚洲传入,经波兰到达西欧。该病正在导致法国白蜡树大量死亡,自2010年首次出现以来传播非常迅速(图12)。
因此,森林树种的衰退是一个复杂现象,与众多气候因素、非生物因素、生物因素和人为因素之间的相互作用有关。每个具体情况仍然是独特的(涉及树种、分布区位置、林学历史、土壤、经营方式等)。没有一种通用的解决方案,但适当的森林经营方法可以显著提高大多数林分对当前主要胁迫的抗性和恢复力。
6.总结
- 森林的破坏与衰退在人类历史早期就已开始,尤其是在人口定居和系统性开垦土地之后,这导致森林面积大幅减少。
- 工业革命标志着树木和森林开始逐渐衰退。这一变化与人类压力(污染、土地利用方式改变等)的不断增加有关。
- 近几十年来(特别是自1990年代和2000年代以来),随着气候变化的加速,这一现象变得更加明显。树木越来越频繁地遭受长期干旱、热浪和臭氧污染的叠加影响。
- 气候胁迫会削弱树木的自然防御能力,使其更容易受到昆虫和真菌的侵袭。
- 与此同时,在全球变暖和全球化的推动下,生物侵袭变得日益严重和频繁。这种侵袭反过来也使树木对气候胁迫更加脆弱。
- 因此,我们正目睹一个真正的恶性循环:气候因素诱发问题并引发衰退,害虫则加剧并加速这一过程,而反复发生的火灾与干旱则相互放大彼此的负面影响。
注释与参考文献
- 封面图片:森林衰退 [照片来源:Colin Behrens,来自Pixabay]
[1] IPCC, 2021: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Masson-Delmotte, V et al.)]. DOI officiel : https://doi.org/10.1017/9781009157896, Lien direct vers la page IPCC : https://www.ipcc.ch/report/ar6/wg1/
[2] Dodane C., « La forêt ardéchoise : une ressource territoriale à révéler et à gérer, une nouvelle organisation de l’espace », Géoconfluences, octobre 2010. – https://geoconfluences.ens-lyon.fr/doc/territ/FranceMut/FranceMutScient8b.htm – Arnould P, Derioz P & Hotyat M (1995). Chapitre 3 : La forêt, pp 53-72. Dans Brunet R & Auriac F. Atlas de France, vol. 6 : Milieux et Ressources, Paris. GIP Reclus/ La Documentation Française. Chiffres 2020 : INSEE et Inventaire forestier de l’IGN. Adaptation : Jean-Benoît Bouron, Géoconfluences, 2021, 2026.
[3] FAO. 2020. Évaluation des ressources forestières mondiales 2020 – Principaux résultats. Rome. https://doi.org/10.4060/ca8753fr
[4] A côté des tempêtes Lothar et Martin en décembre 1999 qui ont dévasté près de 1 million d’ha de forêts françaises, les tempêtes Nils et Pedro de février 2026 ont couché les conifères (pins et épicéas) du massif d’Espinouse dans l’Hérault.
[5] Hammond WM et al. (2022). Global field observations of tree die-off reveal hotter-drought fingerprint for Earth’s forests. Nature Communications, 13, 1761.
[6] Allen CD et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management, 259, 660-684.
[7] Vennetier M. (2006). Pourquoi les forêts brûlent-t-elles ? Edition Le Pommier, 64p.
[8] Claessens H (2016). Quelques considérations pour adapter nos forêts aux changements climatiques. Silva Belgica, Janvier-Février 2016, 20-29.
[9] Mirabel A, Derroire G, Sist P & Traissac S (2025). Comment une forêt peut-elle émettre plus de CO2 qu’elle n’en capture ? The Conversation, 2/10/2025.
[10] Brown F, Sitch S, Folberth G, Cheesman A, Mercado L & Barningham S (2024). Fire activity and drought increase ozone-plant damage to the Amazon rainforest, Physical Sciences. https://doi.org/10.21203/rs.3.rs-4802855/v1.
[11] Wittig VE, Ainsworth EA, Naidu SL, Karnosky DF & Long SP (2009). Quantifying the impact of current and future tropospheric ozone on tree biomass, growth, physiology and biochemistry: a quantitative meta-analysis. Global Change Biology, 15, 396–424, doi: 10.1111/j.1365-2486.2008.01774.x
[12] Dizengremel P (2021). L’ozone et les plantes, histoire d’une relation toxique. The Conversation, 2 mai 2021
[13] Ferretti M et al. (2024). The fingerprint of tropospheric ozone on broadleaved forest vegetation in Europe. Ecological Indicators, 158, 111486.
[14] De Marco A et al. (2022). Strategic roadmap to assess forest vulnerability under air pollution and climate change. Global Change Biology, 28, 5062-5085. DOI: 10.1111/gcb.16278
[15] Bréda N, Huc R, Granier A & Dreyer E (2006). Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Annals of Forest Science, 63, 625-644.https://doi.org/10.1051/forest:2006042
[16] Vitasse Y, Wohlgemuth T & Rigling A (2023). Les forêts face aux sécheresses et canicules : causes de dépérissements, facteurs aggravants et différences de sensibilité entre les espèces. Revue forestière française, 74, 121-132. https://doi.org/10.20870/revforfr.2023.7586
[17] Lu R, Williams L.J, Trouvé R et al. (2026). Pervasive increase in tree mortality across the Australian continent. Nature Plants 12, 62–73. https://doi.org/10.1038/s41477-025-02188-2
[18] Taccoen A., Piedallu C., Seynave I., Gégout-Petit A. & Gégout J. C. (2022). Climate change-induced background tree mortality is exacerbated towards the warm limits of the species ranges. Annals of Forest Science, 79(1).
[19] Lemaire J, Vennetier M, Prévosto B, Cailleret M, & Marty P. (2024). Le dépérissement du pin sylvestre dans le sud-est de la France : des interactions complexes entre climat, topographie, sol et parasite. Forêt méditerranéenne 45, 137-152.
[20] Vennetier M. (2023). Les forêts méditerranéennes face au changement climatique. Quelles leçons pour celles de la Sainte Baume ? Forêt Méditerranéenne 44, 93-108.
[21] Le Cornet L. (2025). Adaptation du massif des Maures au changement climatique. Mémoire de fin d’études, ISTOM, pp81
[22] https://www.notre-environnement.gouv.fr/actualites/breves/article/l-inrae-alerte-sur-les-consequences-du-dereglement-climatique-sur-les-feux-de
[23] Lemaire J (2025). Comprendre et modéliser le risque de dépérissement d’essences forestières dans un contexte de changement climatique. Thèse de doctorat, Aix-Marseille Université – INRAE, 351 p.
[24] Stark R & Cobb F (1969). Smog injury, root diseases and bark beetle damage in Ponderosa pine. California Agriculture, 23 (9), 13-15.
[25] Ochoa-Hueso R et al. (2017). Ecological impacts of atmospheric pollution and interactions with climate change in terrestrial ecosystems of the Mediterranean Basin: Current research and future directions. Environmental Pollution, 227, 194-206.
[26] Lemaire J, Vennetier M, Prévosto B, & Cailleret M. (2025). Multiscale modelling of european beech decline: the role of long-term climate deviations and local environmental factors. European J. Forest Research, 144, 421-441. https://doi.org/10.1007/s10342-025-01767-4.
[27] Saintonge F-X, Gillette M, Blaser S, Queloz V & Leroy Q. (2022). Situation et gestion de la crise liée aux scolytes de l’Épicéa commun fin 2021 dans l’est de la France, en Suisse et en Wallonie. Revue forestière française 73, 619-641. https://doi.org/10.20870/revforfr.2021.7201
[28] Lemaire J, Vennetier M, Prévosto B, & Cailleret M. (2022). Interactive effects of abiotic factors and biotic agents on Scots pine dieback: A multivariate modeling approach in southeast France. Forest Ecology and Management 526, 120543.
[29] Foest J, Bogdziewicz M, Pesendorfer M, Ascoli D, Cutini A, et al. (2024). Widespread breakdown in masting in European beech due to rising summer temperatures. Global Change Biology, 30, e17307.
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: DIZENGREMEL Pierre, VENNETIER Michel (2026年5月24日), 森林与全球环境变化, 环境百科全书,咨询于 2026年7月27日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/forests-global-environmental-change/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。