




为什么虎蚊有如此强的入侵性?

入侵物种究竟来自何方?它们究竟是如何被引入的?全球贸易不断增长,让外来昆虫种类的引入频率实现了爆炸性增长。然而,外来物种是否能够成功实现入侵取决于诸多因素,尤其受入侵物种在生态学和遗传学特征的影响。诚然,绝大多数物种的入侵都以失败告终,但受到全球化的影响,在适当的时间和地点引入更具入侵性物种的可能性大大提高。针对入侵种群进行基因分析,能够获取一系列有价值的信息,从而揭示生物在全球范围和局部地区的引入和扩张的发生机制。
1. 物种生物学
入侵物种是指其种群从一个区域迁移至另一个相隔较远的区域(引入物种种群),并且能够实现繁殖(建立物种种群)并扩展其地理分布(实现物种种群入侵)的物种。这一入侵过程的机制与生态学、健康状况、经济状况这些因素无关。不过,生物入侵被认为是生物多样性变化的主要原因之一[1]。
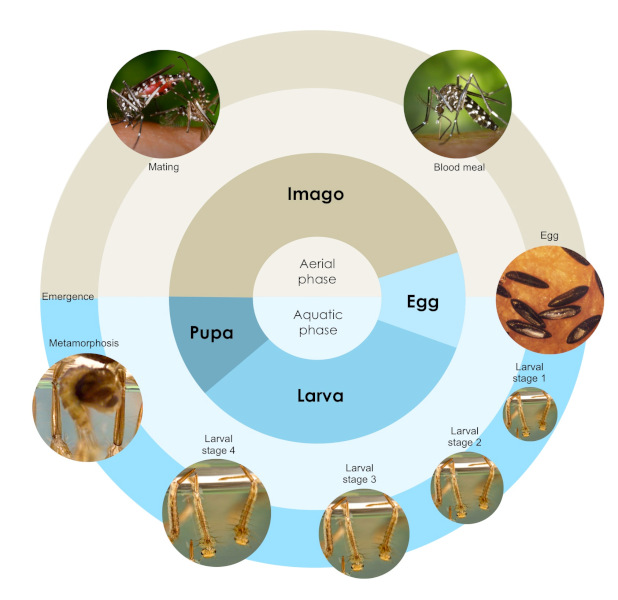
[图片来源:©斯蒂芬妮·夏尔巴(Stéphanie Sherpa),见参考文献5;图片来源:疾病预防与控制中心/亨利·D·普拉特(Henry D. Pratt)/詹姆斯·盖塔尼(James Gathany),由公共卫生图像库提供](Aerial phase 陆生阶段;Aquatic phase 水生阶段;Imago 成虫;Egg 卵;Larva 幼虫;Pupa 蛹;Blood meal 血餐;stage 龄期;Metamorphosis 变态;Emergence 羽化;Mating 交配)
在绝大多数的入侵物种中,昆虫的种类仅次于植物,尤其在北美洲及欧洲地区更是如此[2]。昆虫入侵会给公共健康造成负面的影响,因为昆虫是许多传染病的传播媒介。在这些昆虫中,亚洲虎蚊—学名白纹伊蚊(Aedes albopictus)—会将多种病毒传播给人类,包括登革热病毒、基肯孔雅热病毒和寨卡病毒。起源于东南亚的亚洲虎蚊现如今已经是全球范围内分布最为广泛的蚊种之一,并且被评为100种最具入侵性的物种之一[3]。
1.1. 生命周期
虎蚊的生活史包括4个阶段:卵、幼虫、蛹、成虫(成虫)。根据4个阶段所处的环境不同还可分为2个时期(图1)。其一是水生阶段,包括卵、幼虫、蛹3个阶段;其二是成虫阶段所处的陆生阶段[4]。雄蚊与雌蚊都以花蜜为食,但雌蚊需要吸食血液才能够为卵发育提供所需的营养。虎蚊的卵会产在临近水面的固体表面,只要这一固体表面被水覆盖,蚊卵就能继续发育,而且蚊卵对于干旱环境有一定的耐受力。
1.2. 生态学

最初,虎蚊是一种栖息于森林中的生物,但如今它们已经适应了城郊和乡村环境。虎蚊幼虫可以在不同的环境中完成发育(图2):
- 自然形成的容器,比如树桩或者观赏凤梨;
- 人造容器,比如废旧轮胎、废弃空罐、花盆碟、雨水收集器。
虎蚊在其亚洲原生地往往分布在各种不同的环境中:从东南亚的热带地区(印度尼西亚群岛大部、马来半岛)到中国东北和日本的温带地区都有分布,并且从中南半岛的亚热带地区到印度的西部也都有虎蚊分布(图3)[5]。
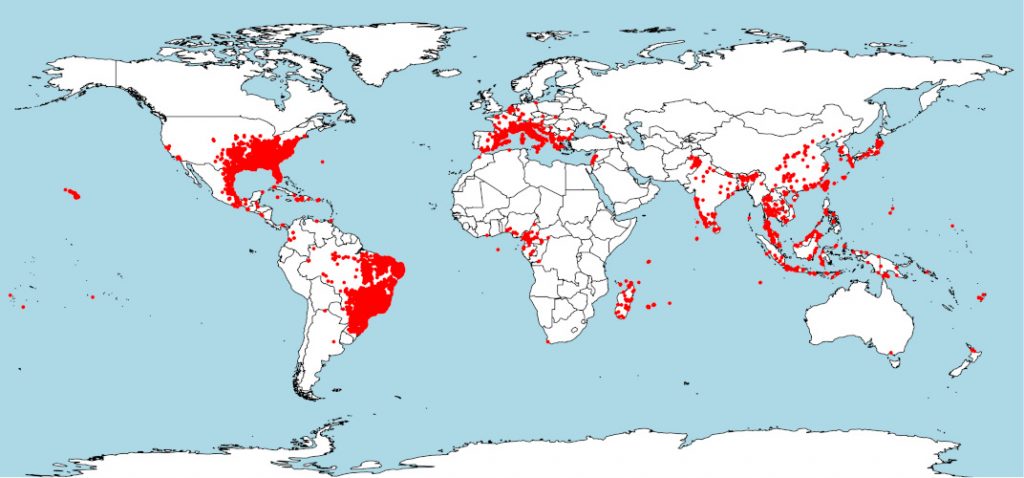
从虎蚊种群的来源地分布来看,其种群丰度呈季节动态变化。在热带地区,虎蚊种群全年都处于活跃状态,但在温带地区,冬季的气候条件便不适合虎蚊生存。不过,虎蚊会产出具有抗寒性的卵,这种卵被称为滞育卵[6]。
2. 入侵种群究竟来自何方?
由于商品贸易、货物和旅客运输业务的发展,外来物种的引入速度在最近的几十年明显加快[2]。
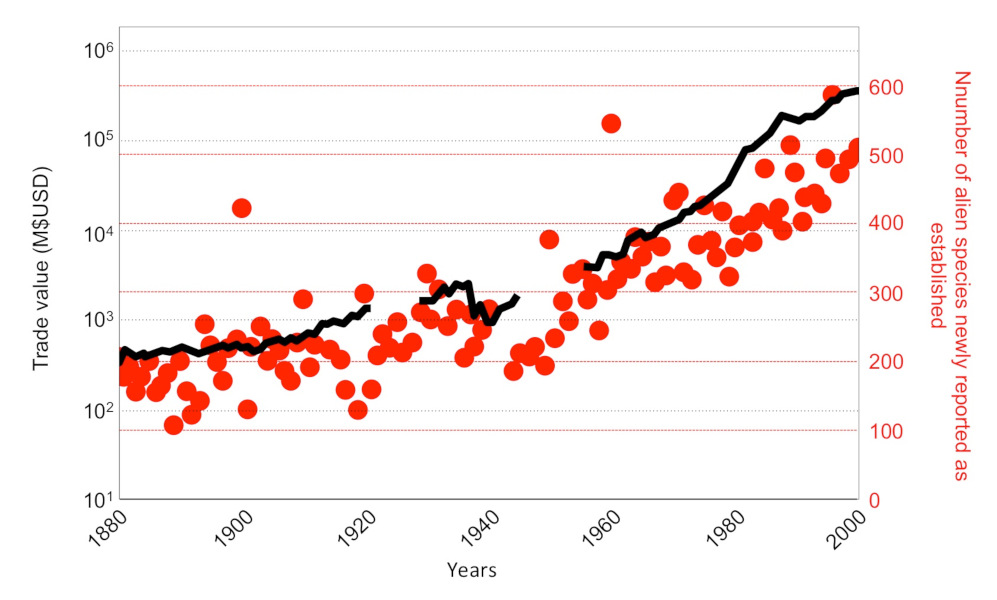
虽然大多数入侵性昆虫在亚洲有各自的原生地,但入侵种群并不总是从它原本的栖息地被直接引入,也有可能是由其他引入的种群带入的,后者被称为“桥头堡”种群。重建外来物种定殖路线,需要依靠遗传研究手段和物种(出现)检测的时空数据,以此为基础描述物种的入侵路线:出发地和目的地、阶段数目、运输方法、引入频率和次数[7]。了解该物种被引入的原因十分必要,可以预防未来的生物入侵[8]。
全球商品贸易的快速增长增加了虎蚊的引入次数。虎蚊的传播可能始于19世纪末,但近几十年来,虎蚊迅速扩展到除南极洲以外的所有大陆,且该物种自20世纪70年代末就已经在欧洲存在[9]。
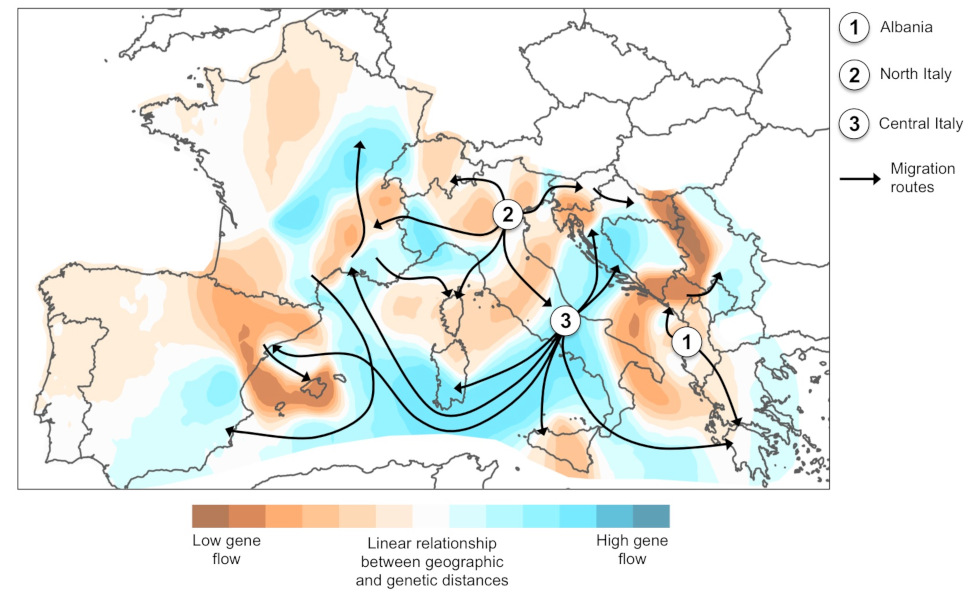
通过对欧洲虎蚊种群进行遗传分析,其结果表明,欧洲的虎蚊种群存在3条相互独立的引入路线:阿尔巴尼亚的虎蚊种群来自中国[10],意大利北部的虎蚊种群来自美国,而意大利中部虎蚊种群的变异则是由意大利北部虎蚊种群与从中国引入的种群杂交而来。全球范围内虎蚊种群的迁移模式表明,当某类虎蚊首次引入某国时,该国彼时的国际贸易伙伴恰好也是该类虎蚊种群的来源地。
尽管每种生物的入侵都有其独有的历程,但对于其他昆虫,诸如亚洲瓢虫[11]和斑翅果蝇[12]之类的入侵物种来说,其入侵路径也存在相似的地理特点。来自亚洲的入侵物种被美国的“桥头堡”种群传播到欧洲,这一过程也显示出了人类的交通运输网络在生物入侵过程中所起的主导作用。
3. 虎蚊在欧洲的迅速扩张
3.1. 种群来源
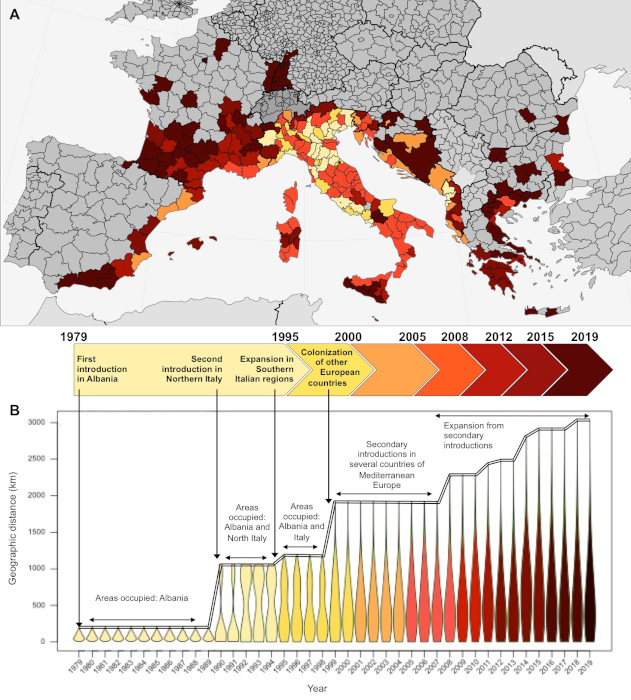
自1979年首次引入阿尔巴尼亚以来,虎蚊于1990年代初被第二次引入了意大利北部[5]。直到20世纪初,虎蚊的地理分布仍然相对局限于这些区域,但在随后不到30年的时间里,虎蚊逐渐侵入地中海沿岸欧洲的所有地区,且每年逐渐向远端扩展(图5)。
通过对虎蚊在欧洲分布的代表性样本进行分析,我们可以精准复原虎蚊在欧洲的扩张历史。最初引入欧洲的三个种群(阿尔巴尼亚、意大利北部、意大利中部)构成了传播中心(图6)。虎蚊种群的迁徙路线反映了人类交通运输的地理足迹:虎蚊种群间的基因交流与地中海盆地的地缘政治背景相关。
3.2. 引入的次数
种群内的遗传多样性决定了种群对环境变化的潜在反应能力以及种群可存续时间[13]。在生物入侵的情况下,入侵物种种群的遗传多样性取决于其来源种群的遗传多样性、引入的个体数量和入侵事件发生的频率[14]。一般来说,刚刚被引入一个新地区的种群个体数量很少,并且只携带了来源种群的一部分基因。这种遗传多样性的匮乏导致个体间高度近亲繁殖,从而增加种群灭绝的风险。
多次引入新种群是提高种群遗传多样性的关键因素之一[15]。这样可以使两组遗传不同的种群个体之间实现交流,不同种群间个体进行交配,最终形成新的基因组合。例如,在意大利中部建立的虎蚊种群是由2个源种群杂交而成,新种群的遗传多样性水平高于其源种群在其栖息的水平[10]。此外,欧洲虎蚊种群之间的高连通性也可能有利于虎蚊在种群扩张进程中维持较高的遗传多样性,从而帮助其在欧洲境内快速扩张(图5)。
4. 易于运输且适应温带气候的物种
贸易决定了虎蚊种群的引入来源以及引入的次数,但唯独不能解释虎蚊为什么能够如此成功地实现入侵。虎蚊的各种生物学及生态学特征在其入侵并建立种群的过程中也起了重要作用。
4.1. 虎蚊引入的可能性
生物为适应人类对其原有栖息地的改造可能会增加它们进入其他区域开辟新的栖息地的可能性,因为有迹象表明,在人类经常活动的地区,生物种群也更容易发生迁移[16]。在虎蚊的各种繁殖场所中,废旧轮胎是其理想的产卵地点。因为虎蚊卵具有抗旱性,可以存活数月,并且能被长途运输。因此,轮胎贸易的大力发展能够使虎蚊卵广泛传播,从而让虎蚊在全球范围内进行扩张。
4.2. 虎蚊建立种群的可能性
虎蚊在温带地区的分布与以下几大气候条件阈值有关[17]。
- 虎蚊卵在冬季最冷月份的存活温度平均高于0°C;
- 维持虎蚊成虫活动的年平均气温为11℃;
- 虎蚊的产卵地最低年降水量须维持在500 mm。
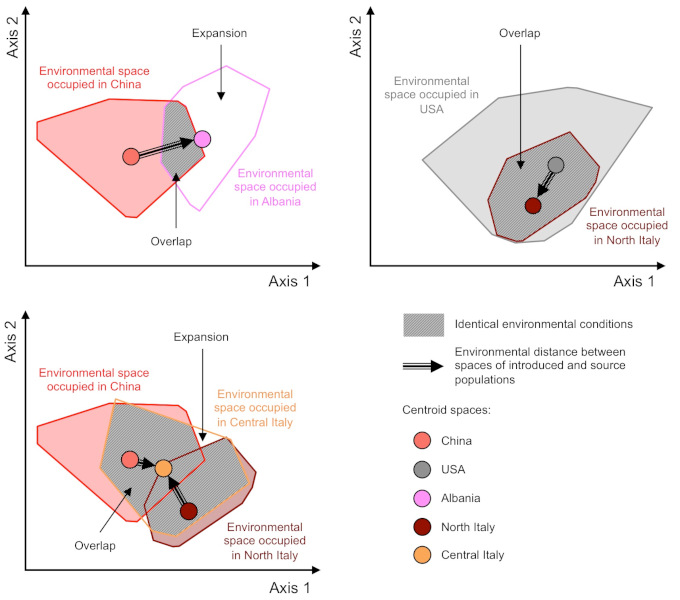
与许多昆虫一样,虎蚊在冬季寒冷环境中的存活依赖于虎蚊卵的滞育能力。这种生理性适应已经出现在亚洲温带地区的虎蚊种群中,那里是阿尔巴尼亚和意大利中部地区虎蚊的来源,而冬季同样寒冷的美国是意大利北部地区虎蚊的来源[5][18][19]。因此,针对欧洲的生存环境,虎蚊已经有了预先的适应。此外,虎蚊种群引入欧洲后所处的环境条件与在其来源地区的环境条件相似[20],这也解释了为什么虎蚊种群在欧洲得以迅速建立(图7)。这些环境条件包括气候条件(温度、降雨量)和人为条件(栖息地的城市化和人工化)。
5. 公路运输造成的虎蚊被动传播
虎蚊被引入后在大陆内扩散,主要原因是虫卵通过公路被动运输,尤其是通过废旧轮胎运输[21]。的确,轮胎存放区发现了大量的虎蚊。然而,最近的研究也强调,日常的汽车运输也会帮助虎蚊成虫扩散[22]。
虎蚊种群的扩散[23]与其基因流动之间存在直接的因果关系。因此,景观遗传学[24]旨在寻找能够促进虎蚊种群间交流的地理和景观因素。与在全球范围内重建虎蚊的入侵路线不同,景观遗传学必须在局部范围内研究虎蚊种群间的连通性。有人类协助的远距离扩散以“跳跃式传播”为特征,而虎蚊种群扩散的前沿地带则是绝佳的天然实验室,有助于研究影响种群连通性的景观因素。
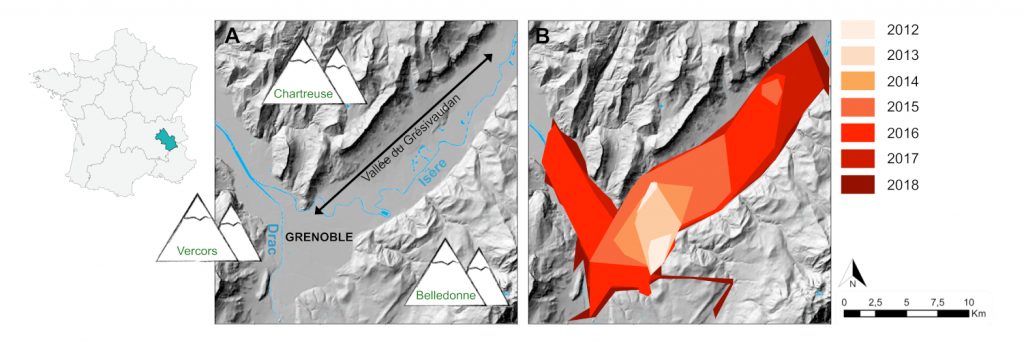
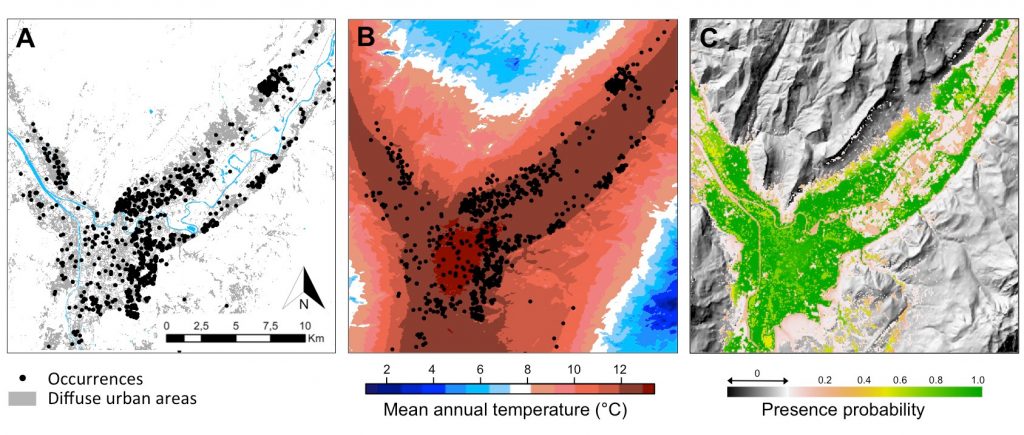
在近期被虎蚊入侵的地区中,格勒诺布尔地区(自2012年以来一直存在虎蚊)是绝佳研究区域,能够帮助了解入侵物种扩张动态。格勒诺布尔地区的虎蚊种群仅可能沿韦科尔、察吐士和贝尔多内地块之间的3个山谷两侧进行扩张(图8)。
在格勒诺布尔地区的景观尺度上,使用不同类型的栖息地(开放栖息地、森林、密集城市地区、住宅城市地区、河流、道路网络)建立模型,模拟影响种群基因多样性的因素,结果表明,沿道路轴线被动扩散的虎蚊与在格勒诺布尔地区两侧远距离地区的虎蚊种群之间存在极强的联系[25](图9)。
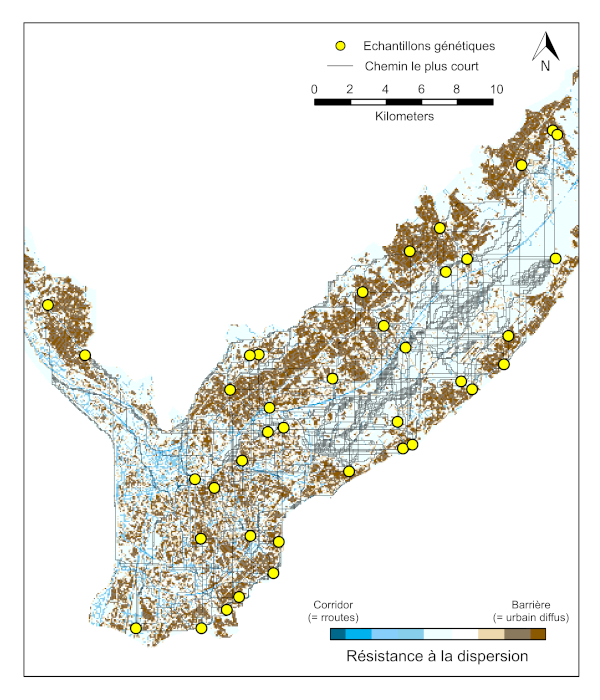
根据捕获再捕获率的估计,虎蚊的自然扩散能力很差,平均每代大约扩散200米[26]。虎蚊大约每3周繁育一代[27],格勒诺布尔地区的环境条件足以使虎蚊从5月一直繁殖到10月,理论上,该地区的虎蚊每年最多可完成7代的繁育。因此,2013年至2017年期间,虎蚊种群中的个体的自然扩散被限制在6 km的半径范围内。由于已确定传播路径两侧基因相近的种群之间的地理距离(图9)可达25 km,本研究证实,相距较远的虎蚊种群之间之所以存在连通性,其主要原因是沿公路的人类交通运输。
6. 随着城市化进程种群的自然快速扩张
在格勒诺布尔地区,虎蚊能够生存取决于(图10):
- 年平均最低温度11°C,保证夏季温度适于成虫保持活跃,冬季温度能让卵存活。该温度阈值对应海拔800m等高线。
- 虎蚊在城市内的栖息地十分分散,通常为带有花园的住宅区,这类区域有大量适宜产卵的地点(花盆碟、集水器)和人类(可充当雌性虎蚊的血餐),帮助虎蚊完成繁殖过程。
尽管在生物入侵的情况下常常被忽视,但自然扩散也可能在小空间尺度上促进种群的扩张。罗纳阿尔卑斯大区蚊虫控制机构(EID)建立了一种扩散模型,利用所收集的物种时空数据以及生态建模(图10),以确定合适栖息地和物种的生物特征(每年的世代数、每代成虫的最大扩散距离),这种模型可区分自然扩散兼容的扩散事件与必须通过被动方式发生的(距离过长)扩散事件。通过比较虎蚊引入以来的连续几年的模型发现,70%到80%的新的年度检测结果可归因于自然扩散(图11)。

这种自然扩散的优势证实了种群遗传分化的模式,公园、私人花园和墓地等区域在城市中较为分散,被认为阻碍了虎蚊种群扩散(图9)。相距数百米到2公里的分散城市区域之间的种群基因相似性较低。之所以扩散率低,是因为这样一个事实,即分散的城市区域为虎蚊提供了充足的有利栖息地,具有相对较多的产卵和繁育地点(用于饲育和繁殖)和植物区(用于成虫休养生息)。
格勒诺布尔地区有大量适宜虎蚊生存的连续栖息地,在虎蚊进入该地几年后,虎蚊几乎在所有居民区都迅速定居下来。人口增长,人口密集城市边缘地带居民区极具吸引力,这都逐渐导致城市居住空间扩大,开放栖息地(如草地或农田)也被取代,这种趋势为虎蚊种群的建立和扩张提供了新的适宜栖息地。土地利用形式的变化和人类对生态系统施加的压力促进了入侵物种种群的建立和加速扩张。
7. 结论
- 对入侵种群的遗传分析可以在全球范围内和局部景观范围内追踪其传播历史(入侵、建立、扩张)。
- 引入的次数和种群的来源地决定了其在当地建立新种群的能力(遗传多样性和适应性特征)并受人类交通运输网络的制约。
- 人类通过公路辅助虎蚊长距离扩散,导致虎蚊种群间频繁交流,有利于虎蚊种群的长期生存。
- 景观尺度上的自然扩散在虎蚊种群空间扩张中的作用往往被低估,种群扩散程度往往取决于适宜栖息地的数量。
- 这些虎蚊栖息地的范围和可用性直接受土地利用方式变化的影响。
参考资料及说明
封面照片:虎蚊。[图片来源:詹姆斯·加萨尼(James Gathany)CDC/公共领域]
[1] Simberloff, D. et al. (2013). Impacts of biological invasions: what’s what and the way forward. Trends in Ecology and Evolution, 28(1), 58-66.
[2] Seebens, H. et al. (2017). No saturation in the accumulation of alien species worldwide. Nature Communications, 8(1), 1-9.
[3] S. Lowe, M. et al. (2007).从全球入侵物种数据库中选择。入侵物种专家组(ISSG).
[4] 下莱茵河蚊子联盟。虎蚊-生物学(法语)。
[5] Sherpa, S. (2019).欧洲虎蚊种群的集群历史和入侵成功的决定因素。博士论文,格勒诺布尔阿尔卑斯大学。
[6] 滞育是发育的暂时停止,具有可选性,抑或具有强制性,通常由环境条件(温度和光周期)决定。
[7] Estoup, A., & Guillemaud, T. (2010). Reconstructing routes of invasion using genetic data: why, how and so what? Molecular Ecology, 19(19), 4113-4130.
[8] Estoup, A. & Guillemaud, T. (1999).追溯入侵以更好地打击入侵。 Pour la Science (法语)
[9] 欧洲疾病预防和控制中心 (ECD)。白纹伊蚊。
[10] Sherpa, S. et al. (2019). Unraveling the invasion history of the Asian tiger mosquito in Europe. Molecular Ecology, 28(9),2360-2377.
[11] 国家农艺研究所(INRA)(2010年)。追踪了亚洲瓢虫Harmonia axyridis的入侵路线。 (法语)
[12] Fraimout, A. et al. (2017). Deciphering the routes of invasion of Drosophila suzukii by means of ABC random forest. Molecular Biology and Evolution, 34(4), 980-996.
[13] Bock, D. G. et al. 2015. What we still don’t know about invasion genetics. Molecular Ecology, 24(9), 2277-2297.
[14] Simberloff, D. 2009. The role of propagule pressure in biological invasions. Annual Review of Ecology, Evolution, and Systematics, 40, 81-102.
[15] Dlugosch, K. M. & Parker, I. M. (2008). Founding events in species invasions: genetic variation, adaptive evolution, and the role of multiple introductions. Molecular Ecology, 17(1), 431-449.
[16] Hufbauer, R. A. et al. 2012. Anthropogenically induced adaptation to invade (AIAI): contemporary adaptation to human-altered habitats within the native range can promote invasions. Evolutionary Applications, 5(1), 89-101.
[17] Fischer, D. (2014). Climatic suitability of Aedes albopictus in Europe referring to climate change projections: comparison of mechanistic and correlative niche modelling approaches. Eurosurveillance, 19, 20696.
[18] Armbruster, P. A. (2016). Photoperiodic diapause and the establishment of Aedes albopictus (Diptera: Culicidae) in North America. Journal of Medical Entomology, 53(5), 1013-1023.
[19] Sherpa, S. et al. (2019). Cold adaptation in the Asian tiger mosquito’s native range precedes its invasion success in temperate regions. Evolution, 73(9), 1793-1808.
[20] Sherpa, S. et al. (2019). Predicting the success of an invader: Niche shift versus niche conservatism. Ecology and Evolution,9(22), 12658-12675.
[21] Scholte, E. J. & Schaffner, F. (2007). Waiting for the tiger: establishment and spread of the Aedes albopictus mosquito in Europe. In Emerging pests and vector-borne diseases in Europe, Eds. Takken W., & Knols, B. G. J., Wageningen Academic Publishers. pp. 241-260.
[22] Eritja, R. et al. (2017). Direct evidence of adult Aedes albopictus dispersal by car. Scientific Reports, 7(1), 1-15.
[23] 生物个体出生地和繁殖地之间的平均距离。自然扩散取决于一个物种的个体在特定环境中的移动能力,而被动扩散则涉及人类对个体移动的干预。
[24] Manel, S. et al. (2003). Landscape genetics: combining landscape ecology and population genetics. Trends in Ecology and Evolution, 18(4), 189-197.
[25] Sherpa, S. et al. (2020) Landscape does matter: disentangling founder effects from natural and human-aided post-introduction dispersal during an ongoing biological invasion. Journal of Animal Ecology, 89(9), 2027-2042.
[26] Marini, F. et al. (2019). Estimating spatio-temporal dynamics of Aedes albopictus dispersal to guide control interventions in case of exotic arboviruses in temperate regions. Scientific Reports,9(1), 1-9.
[27] 一代个体产生新一代个体所需的平均时间。
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: SHERPA Stéphanie (2024年2月23日), 为什么虎蚊有如此强的入侵性?, 环境百科全书,咨询于 2026年7月28日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/%e4%b8%ba%e4%bb%80%e4%b9%88%e8%99%8e%e8%9a%8a%e6%9c%89%e5%a6%82%e6%ad%a4%e5%bc%ba%e7%9a%84%e5%85%a5%e4%be%b5%e6%80%a7%ef%bc%9f/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。


