


Why is the tiger mosquito so invasive?
PDF
Where do invasive species come from? How are they introduced? Increased trade has led to an explosion in the frequency of introduction of alien insects on a global scale. However, many factors are decisive for the success of a biological invasion, in particular the ecological and genetic characteristics of introduced populations. Indeed, most species introductions fail. Globalization increases the likelihood of introducing an invasion-prone species at the right time and place. Genetic analysis of invasive populations provides valuable information on how introductions and expansion are taking place, both on a global scale and on a local landscape scale.
1. Biology of the species
An invasive species is one whose populations have been transported from one region to another geographically distant region (introduced populations), and which once introduced are capable of reproducing (established populations) and expanding their geographic distribution (invasive populations). The mechanisms at work during this invasion process are independent of the notion of impacts, whether ecological, health or economic. Nevertheless, biological invasions are recognized as one of the main causes of biodiversity change [1].
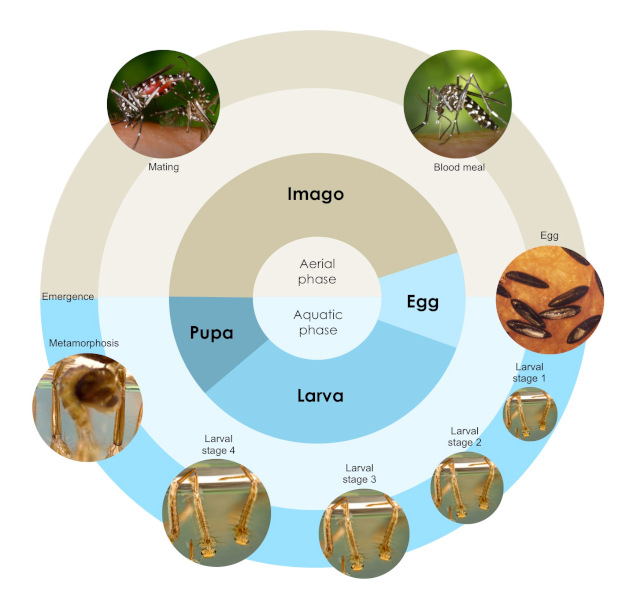
1.1. Life cycle
The tiger mosquito life cycle consists of four stages: egg, larva, pupa, and imago (adult), divided into two phases depending on the environment in which the stages occur (Figure 1). The first phase, aquatic, consists of the egg, larva, and pupa stages, with the adult stage being aerial [4]. Both male and female adult mosquitoes feed on nectar, but the females require a blood meal to provide the nutrients needed for egg development. The eggs are laid on a solid surface close to the water surface, and continue to develop once the surface is covered with water. The eggs are drought resistant.
1.2. Ecology

- natural containers, such as tree stumps, or Bromeliads ;
- artificial containers, such as used tires, abandoned containers, flowerpot saucers, or rainwater catchers.
In its Asian native range, the tiger mosquito occurs in a wide range of environments: from tropical areas of Southeast Asia (large Indonesian islands, Malay Peninsula) to temperate regions of Northeast China and Japan, and from subtropical areas of the Indochinese Peninsula to India further west (Figure 3). [5]
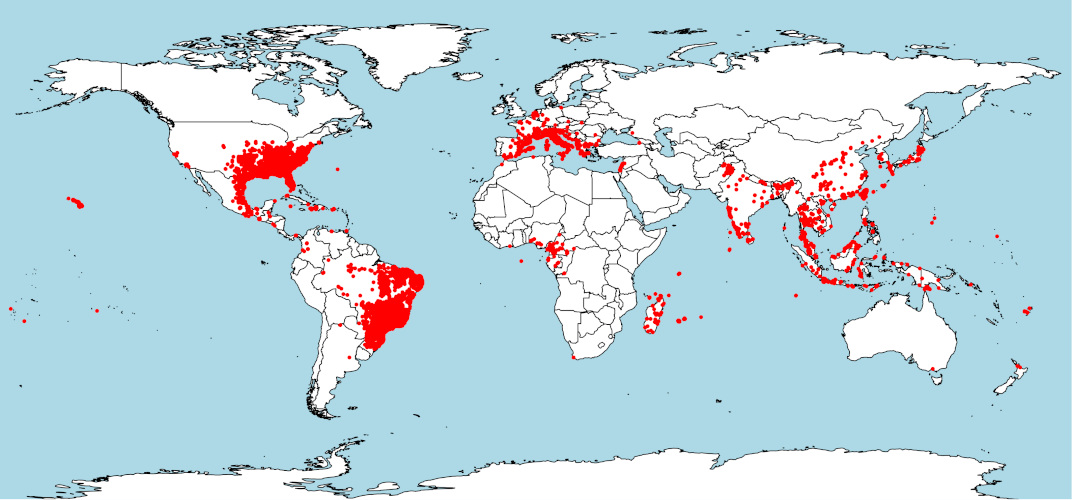
2. Where do invasive populations come from?
The rate of introduction of alien species has increased significantly in recent decades [2], in particular due to increased trade and transport of goods and people (Figure 4).
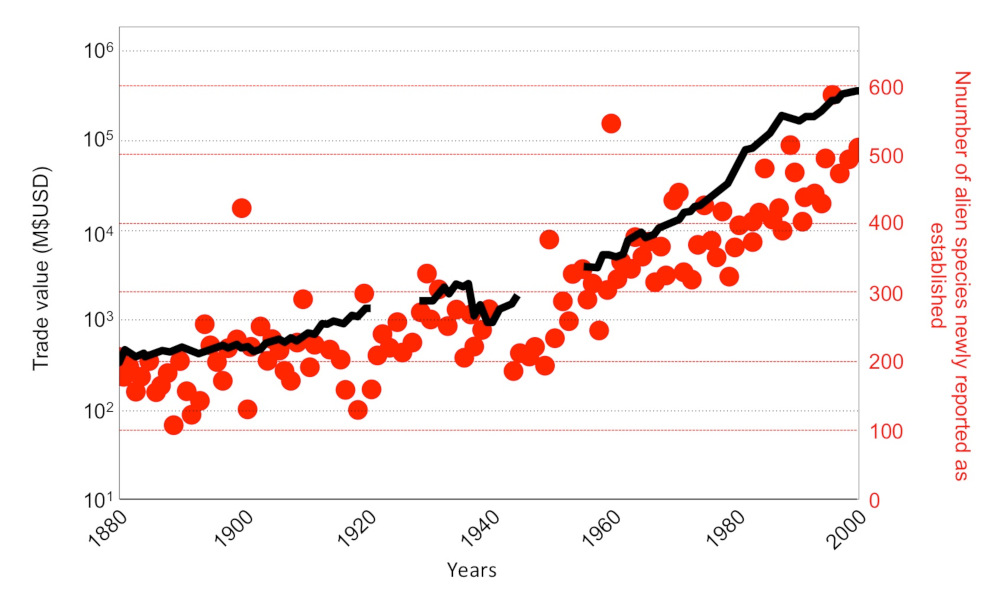
The rapid growth in global merchandise trade has increased the number of tiger mosquito introductions. Its spread probably began in the late 19th century, but its distribution has rapidly spread to all continents except Antarctica in recent decades, particularly in Europe where the species has been present since the late 1970s [9].
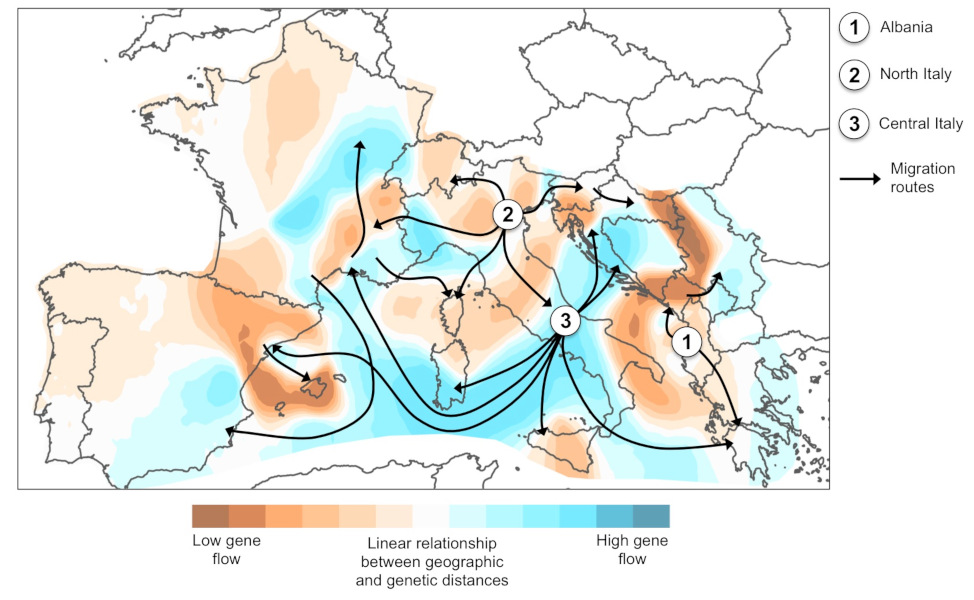
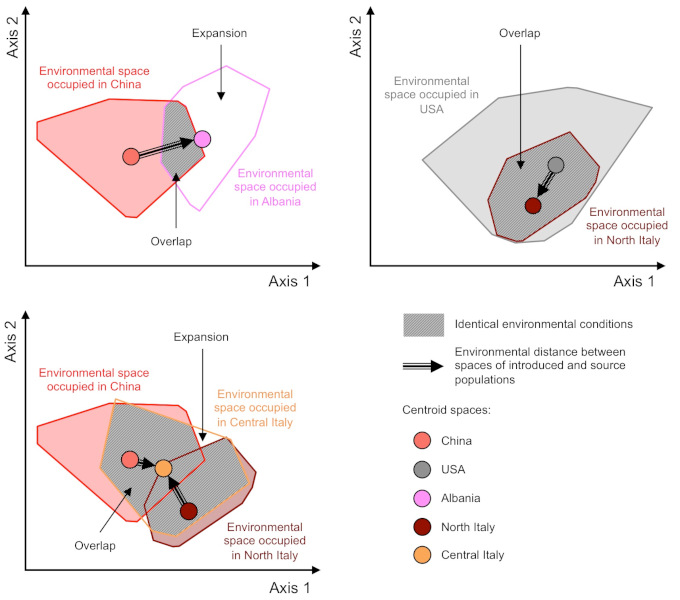
Genetic analysis of European populations shows three independent introductions: in Albania from China, in northern Italy from the USA, and in central Italy, where the genetic variability of the populations is the result of a mixture between those initially present in northern Italy and a new introduction from China [10]. Migration patterns at the global scale demonstrate that the source populations at the origin of the introductions correspond to international trade partners at the time of the first introductions.
Although each biological invasion has its own history, introductions of other insect species, such as the Asian ladybird [11] and the spotted-wing drosophila [12], have followed a similar geographical pattern. The introduction into Europe has taken place from Asia via bridgehead populations in the United States, suggesting a predominant role of human transport networks.
3. Rapid expansion in Europe
3.1. Origin of populations

The analysis of a representative sample of the distribution of the species in Europe has made it possible to reconstruct the precise history of the expansion. The three populations initially introduced in Europe (Albania, Northern Italy, Central Italy) constituted centres of dispersal (Figure 6). The migration routes reflect the geography of human transport: genetic exchanges correlate to the geopolitical context in the Mediterranean basin.
3.2. Number of introductions
Multiple introductions are one of the main factors increasing population genetic diversity [15]. They can bring two genetically distinct groups of individuals into contact, whose mixing leads to the appearance of new genetic combinations. For example, the tiger mosquito populations established in central Italy, resulting from mixing between two source populations, have higher levels of genetic diversity than those recorded in the native range of the species [10]. Furthermore, the high connectivity between introduced populations has probably favoured the maintenance of high levels of genetic diversity within populations on the expansion fronts, contributing to the rapid expansion in Europe (Figure 5).
4. A species predisposed to transport and temperate climates
Trade determines the origin of introduced populations and the number of introductions, but cannot explain alone the success of the tiger mosquito. Different elements of its biology and ecology have also played a role in introduction and establishment success.
4.1. Probability of introduction
The adaptation of organisms to human-modified habitats may increase the likelihood of their introduction into new regions because populations in areas frequented by humans are more likely to be transported [16]. Among the breeding sites used by tiger mosquitoes, used tires are the preferred egg-laying site. Because they are drought resistant, the eggs can survive for several months and can be transported over long distances. Intensified tire trade has thus initiated the global expansion of the species through the transport of eggs.
4.2. Probability of establishment
The distribution of tiger mosquitoes in temperate regions corresponds to several thresholds of climatic conditions [17]:
- an average temperature in the coldest month above 0°C for the winter survival of eggs;
- an average annual temperature of 11°C for the maintenance of adult activity;
- a minimum annual precipitation of 500 mm for the maintenance of egg-laying sites.
As with many insects, the winter survival of the tiger mosquito in cold environments is primarily determined by the ability of the eggs to enter diapause. This physiological adaptation is already present in temperate regions of Asia, where are the sources of the introduction in Albania and central Italy, and in the USA, which is the source of the introduction in northern Italy [5], [18],[19]. Thus, the tiger mosquito was already pre-adapted to settle in Europe. Moreover, the environmental conditions encountered during introduction in Europe were similar to those encountered in the source regions [20], which may explain the rapid establishment of the populations (Figure 7). These environmental constraints include climatic constraints (temperature, rainfall) but also anthropogenic constraints (urbanisation and artificialisation of habitats).
5. Passive dissemination via road transport
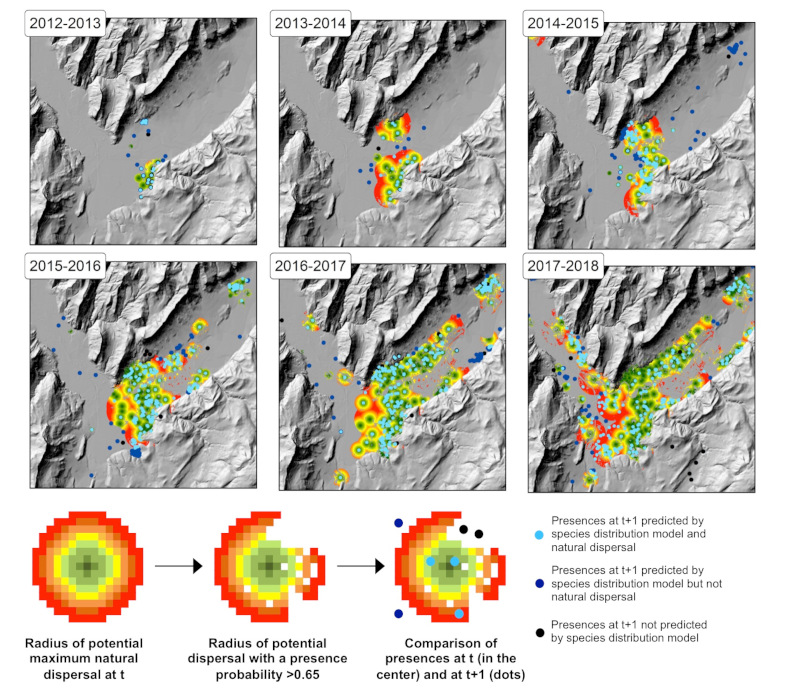
There is a direct causal relationship between dispersal [23] and gene flow between populations. Therefore, landscape genetics [24] seeks to identify geographic and landscape factors that promote population connectivity. In contrast to colonisation routes, that are reconstructed at a global scale, population connectivity must be studied at a local scale. Although human-assisted long-distance dispersal can be characterized by “jumps”, expansion fronts provide excellent natural laboratories for studying landscape factors affecting population connectivity.
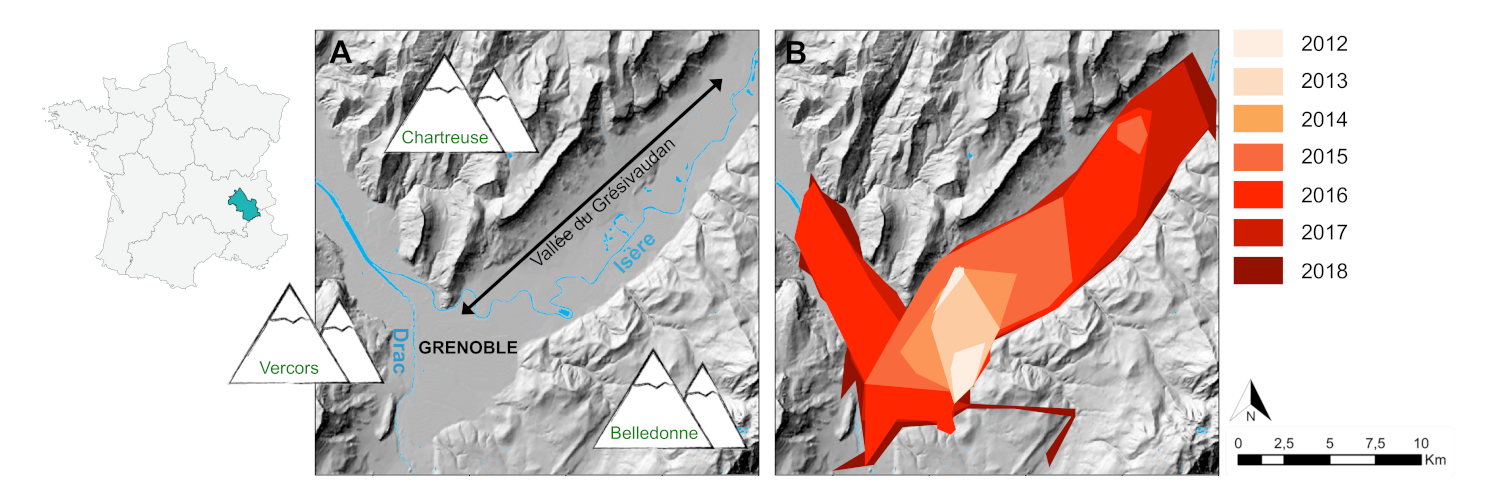
Modelling the factors structuring the genetic variability of populations at the Grenoble landscape scale, using different types of habitats (open habitats, forests, dense urban areas, residential urban areas, rivers, road networks), revealed that passive dispersal of tiger mosquitoes along road axes induces strong connectivity between geographically distant populations on either side of Grenoble [25] (Figure 9).
The tiger mosquito has a low natural dispersal capacity of about 200m per generation on average based on estimations from capture-recapture rates [26]. The generation time [27] is about three weeks in the tiger mosquito. Environmental conditions in the Grenoble region being favourable for reproduction from May to October, the species can theoretically complete up to seven generations per year. Thus, the maximum natural dispersal of individuals between 2013 and 2017 is limited to a radius of 6 km. As the geographical distance between genetically close populations on either side of identified roads (Figure 9) can be up to 25 km, this study confirms that the main factor explaining the connectivity between distant populations in the tiger mosquito is human transport along roads.
6. Rapid natural expansion due to urbanization of the territories
In the Grenoble area, the presence of the tiger mosquito is determined by (Figure 10):
- A minimum mean annual temperature of 11°C, correlated to an optimal temperature during the summer months, when the adults are active, and to a winter temperature allowing the survival of the eggs. This temperature threshold corresponds to the 800m isocline.
- The presence of diffuse urban habitats, corresponding to residential areas with gardens where egg-laying sites (flowerpot cups, water collectors) and humans (blood meal for females) are abundant, allowing the reproductive cycle to be completed.
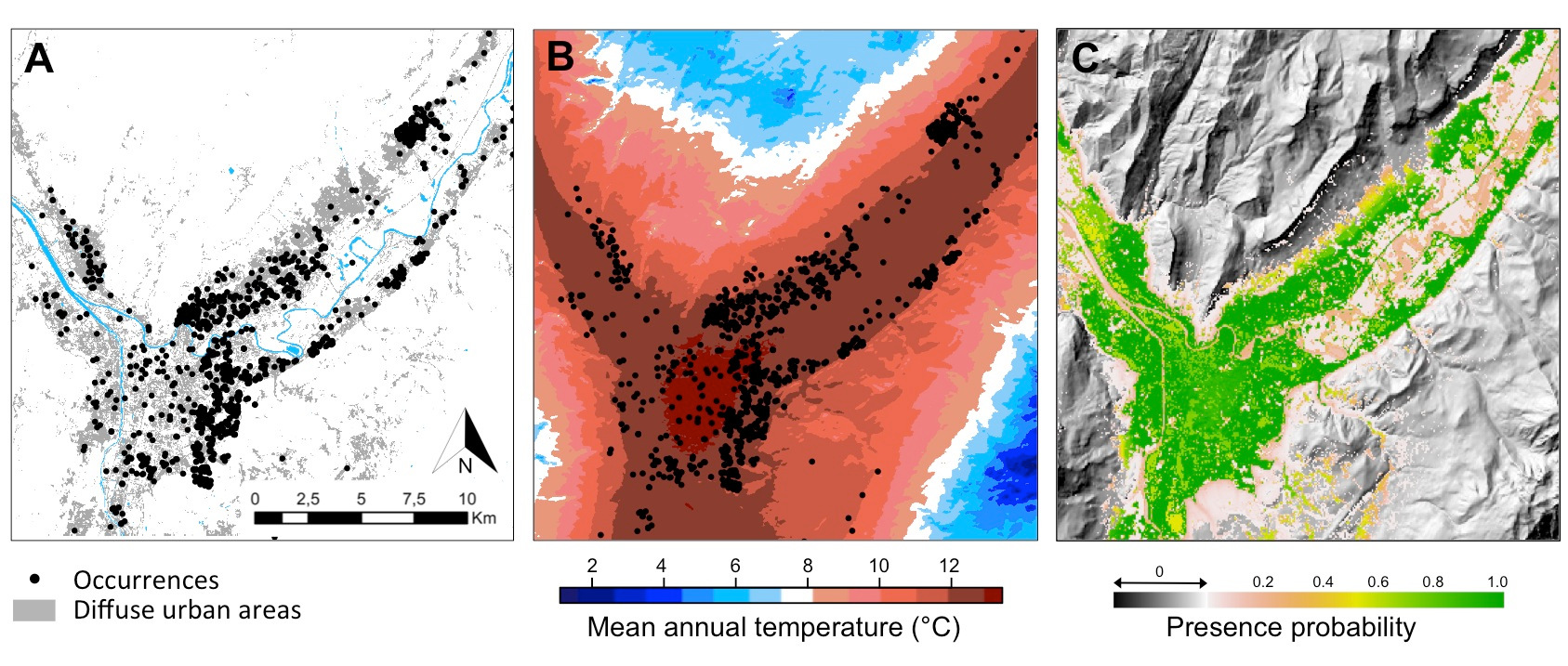
The large proportion of continuous suitable habitats for the tiger mosquito in the Grenoble region promoted the rapid colonisation of almost all residential areas a few years after its introduction. Human population growth and the attractiveness of residential areas on the edges of dense urban areas gradually induced an extension of urban residential spaces and a replacement of open habitats, such as meadows or farmland, providing new suitable habitats for the establishment and expansion of the tiger mosquito. Changes in land use and human pressures on ecosystems promote the establishment and accelerate the expansion of invasive species (See When invasive plants also walk in the fields).
7. Messages to remember
- Genetic analysis of invasive populations makes it possible to trace their history (introduction, establishment, expansion) at the global scale, but also at the local landscape scale.
- The number of introductions and the origin of introduced populations determining their capacity to establish (genetic diversity and adaptive traits) are conditioned by human transport networks.
- Human-assisted long-distance dispersal along motorways, leading to frequent exchanges between populations, favours their long-term persistence.
- The role of natural dispersal at the landscape scale in the spatial expansion of populations is often underestimated and depends on the amount of favourable habitats.
- The extent and availability of these habitats is directly influenced by land use change.
Notes and References
Cover image. Tiger mosquito. [Source: James Gathany, CDC/public domain]
[1] Simberloff, D. et al. (2013). Impacts of biological invasions: what’s what and the way forward. Trends in Ecology and Evolution, 28(1), 58-66.
[2] Seebens, H. et al. (2017). No saturation in the accumulation of alien species worldwide. Nature Communications, 8(1), 1-9.
[3] S. Lowe, M. et al. (2007). 100 of the world’s most harmful invasive alien species. A selection from the Global Invasive Species Database. Invasive Species Specialist Group (ISSG).
[4] Syndicat de Lutte contre les Moustiques du Bas-Rhin. Moustique tigre – Biologie (in French).
[5] Sherpa, S. (2019). History of colonization and determinants of invasive success of tiger mosquito populations in Europe. PhD thesis, Université Grenoble Alpes.
[6] Diapause is a temporary cessation of development, optional or mandatory, and generally determined by environmental conditions (temperature and photoperiod).
[7] Estoup, A., & Guillemaud, T. (2010). Reconstructing routes of invasion using genetic data: why, how and so what? Molecular Ecology, 19(19), 4113-4130.
[8] Estoup, A. & Guillemaud, T. (1999). Retracer l’invasion pour mieux la combattre. Pour la Science (in French)
[9] European Center for Disease Prevention and Control (ECD). Aedes albopictus.
[10] Sherpa, S. et al. (2019). Unraveling the invasion history of the Asian tiger mosquito in Europe. Molecular Ecology, 28(9), 2360-2377.
[11] Institut National de la Recherche Agronomique (INRA) (2010). Les routes d’invasion de la coccinelle asiatique Harmonia axyridis retracées. (in French)
[12] Fraimout, A. et al. (2017). Deciphering the routes of invasion of Drosophila suzukii by means of ABC random forest. Molecular Biology and Evolution, 34(4), 980-996.
[13] Bock, D. G. et al. 2015. What we still don’t know about invasion genetics. Molecular Ecology, 24(9), 2277-2297.
[14] Simberloff, D. 2009. The role of propagule pressure in biological invasions. Annual Review of Ecology, Evolution, and Systematics, 40, 81-102.
[15] Dlugosch, K. M. & Parker, I. M. (2008). Founding events in species invasions: genetic variation, adaptive evolution, and the role of multiple introductions. Molecular Ecology, 17(1), 431-449.
[16] Hufbauer, R. A. et al. 2012. Anthropogenically induced adaptation to invade (AIAI): contemporary adaptation to human-altered habitats within the native range can promote invasions. Evolutionary Applications, 5(1), 89-101.
[17] Fischer, D. (2014). Climatic suitability of Aedes albopictus in Europe referring to climate change projections: comparison of mechanistic and correlative niche modelling approaches. Eurosurveillance, 19, 20696.
[18] Armbruster, P. A. (2016). Photoperiodic diapause and the establishment of Aedes albopictus (Diptera: Culicidae) in North America. Journal of Medical Entomology, 53(5), 1013-1023.
[19] Sherpa, S. et al. (2019). Cold adaptation in the Asian tiger mosquito’s native range precedes its invasion success in temperate regions. Evolution, 73(9), 1793-1808.
[20] Sherpa, S. et al. (2019). Predicting the success of an invader: Niche shift versus niche conservatism. Ecology and Evolution, 9(22), 12658-12675.
[21] Scholte, E. J. & Schaffner, F. (2007). Waiting for the tiger: establishment and spread of the Aedes albopictus mosquito in Europe. In Emerging pests and vector-borne diseases in Europe, Eds. Takken W., & Knols, B. G. J., Wageningen Academic Publishers. pp. 241-260.
[22] Eritja, R. et al. (2017). Direct evidence of adult Aedes albopictus dispersal by car. Scientific Reports, 7(1), 1-15.
[23] Average distance travelled between birth and reproduction of an individual. Natural dispersal depends on the ability of individuals of a species to move in a given environment, whereas passive dispersal involves human intervention in the movement of individuals.
[24] Manel, S. et al. (2003). Landscape genetics: combining landscape ecology and population genetics. Trends in Ecology and Evolution, 18(4), 189-197.
[25] Sherpa, S. et al. (2020) Landscape does matter: disentangling founder effects from natural and human-aided post-introduction dispersal during an ongoing biological invasion. Journal of Animal Ecology, 89(9), 2027-2042.
[26] Marini, F. et al. (2019). Estimating spatio-temporal dynamics of Aedes albopictus dispersal to guide control interventions in case of exotic arboviruses in temperate regions. Scientific Reports, 9(1), 1-9.
[27] The average time it takes for individuals in one generation to produce a new generation of individuals.
The Encyclopedia of the Environment by the Association des Encyclopédies de l'Environnement et de l'Énergie (www.a3e.fr), contractually linked to the University of Grenoble Alpes and Grenoble INP, and sponsored by the French Academy of Sciences.
To cite this article: SHERPA Stéphanie (March 9, 2021), Why is the tiger mosquito so invasive?, Encyclopedia of the Environment, Accessed August 3, 2026 [online ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/en/life/why-tiger-mosquito-invasive/.
The articles in the Encyclopedia of the Environment are made available under the terms of the Creative Commons BY-NC-SA license, which authorizes reproduction subject to: citing the source, not making commercial use of them, sharing identical initial conditions, reproducing at each reuse or distribution the mention of this Creative Commons BY-NC-SA license.


