


Evolution of colour vision in mammals and primates
PDFIn non-mammalian vertebrates, visual and non-visual light reception involves ocular, lateral and extraocular photoreceptor structures respectively: pineal organ, parapineal organ, frontal organ, parietal eye and deep brain photoreceptors. These different organs allow individuals to collect, from a single light source, various information such as photoperiod, light intensity and electromagnetic spectrum. This makes it possible to reduce misinterpretation and refine the measurement of light and therefore the measurement of time. However, in mammalian species, visual and non-visual photoreceptions involve only ocular structures. This particularity could be linked to their evolution since a common nocturnal ancestor appeared 225 million years ago. Nowadays, 63.8% of mammals are nocturnal. However, navigating in darkness requires very sensitive structures that can distinguish minute variations in light intensity or the electromagnetic spectrum. Only the ocular photoreceptor structures are sensitive enough to distinguish these variations. This constraint would have led mammals to lose their extraocular photoreceptor structures. This is also observed in nocturnal non-mammalian species such as geckos that have lost their parietal eyes.
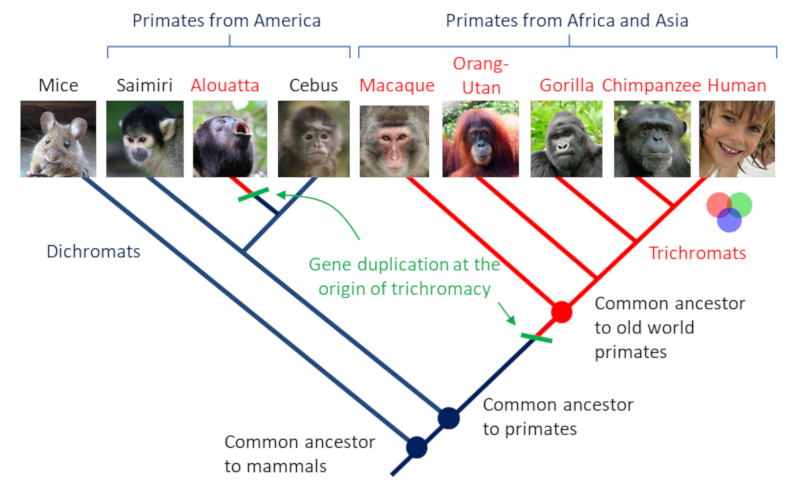
However, some primates are the exception. Indeed, while their common ancestor was dichromate, several species (including humans) have once again acquired the trichromatic vision. Thus, among catarhins (monkeys of the “Old World”) and alouates (South American howler monkeys), there are three genes, called SWS1, MWS and LWS, that code for three types of cones. In platyrhinians (monkeys of the “New World”) there are two genes, one of which is poly-allelic (there are several forms of the same SWS1 gene and three LWS allelic forms) and linked to the X chromosome. In other words, for these species, only heterozygous females, which have two different LWS alleles, have trichromatic vision; homozygous females, which have two identical LWS alleles and males, which have only one X chromosome and only one LWS allele, are dichromatic. The reason for a return to trichromatics? It seems that the trichromatic vision has given these primates a selective advantage in distinguishing ripe red fruits on the green background from vegetation. Indeed, the MWS gene and one of the LWS alleles encode cones that are sensitive to the green color.
References and notes
[1] Adapted from http://acces.ens-lyon.fr/acces/ressources/neurosciences/vision/enseigner/pigments-retiniens-et-evolution/les-pigments-retiniens-des-outils-pour-retracer-l2019evolution