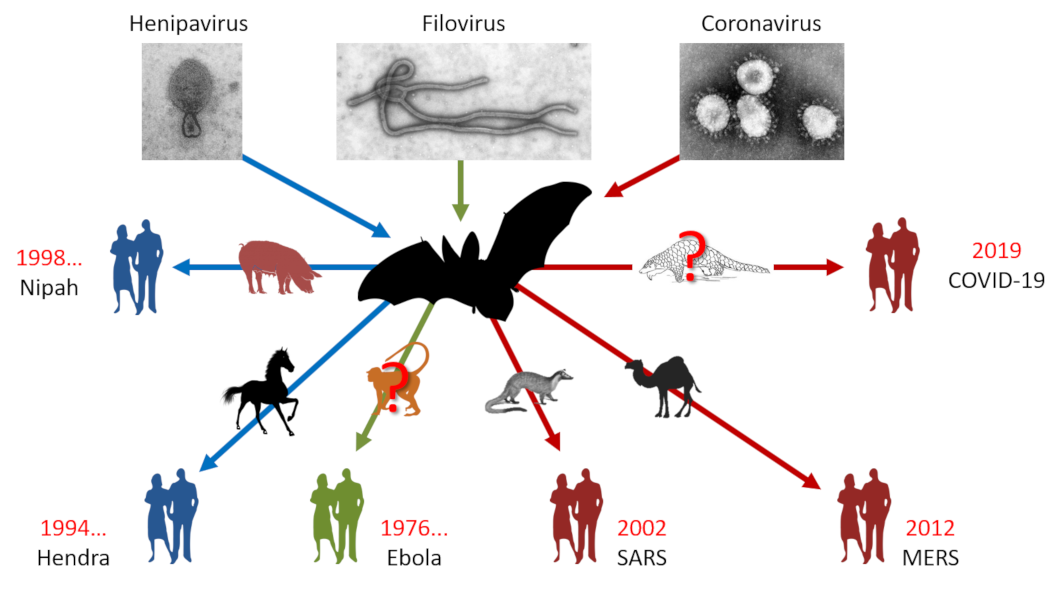
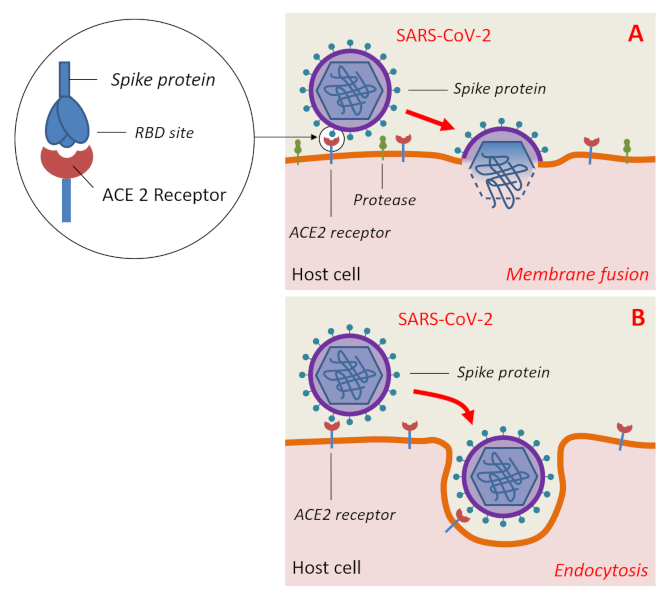
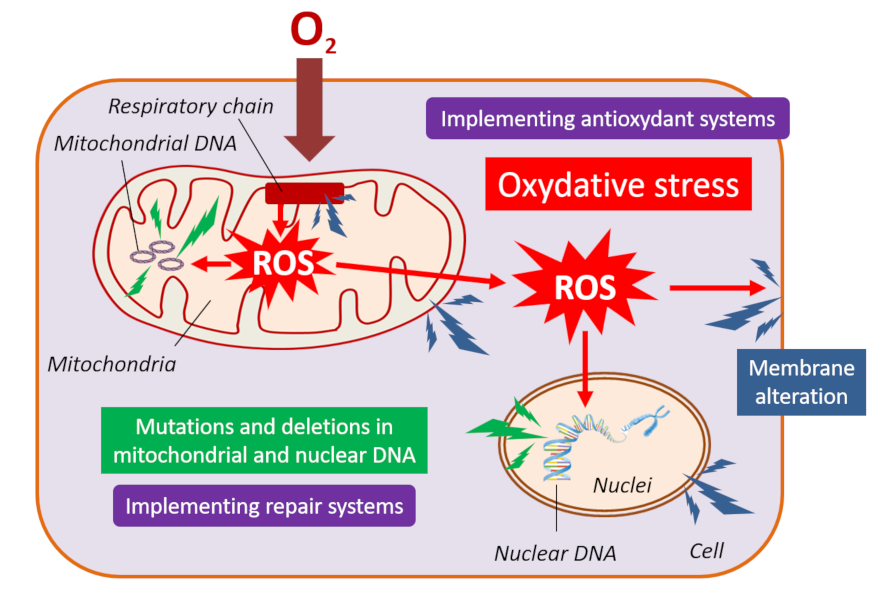
图1. 蝙蝠是很多致病新型病毒的天然宿主。 [来源:亨尼巴病毒属,CSIRO, CC BY 3.0,通过维基共享 /丝状病毒科,图片来源:CDC/辛西娅·戈德史密斯(Cynthia Goldsmith),公共领域,通过维基共享 /冠状病毒科,公共领域] (Henipavirus:亨尼巴病毒属;Filovirus:丝状病毒科;Coronavirus:冠状病毒科;Nipah:尼帕;Hendra:亨德拉;Ebola:埃博拉;SARS:严重急性呼吸综合征;MERS:中东呼吸综合征;COVID-19:2019冠状病毒病)
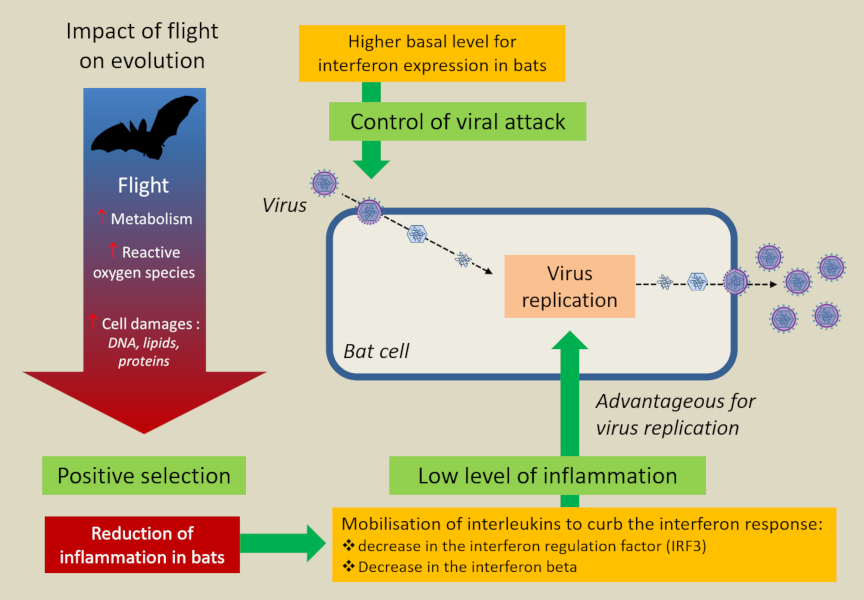
图15. 蝙蝠控制病毒和炎症的独特免疫能力源自飞行。[来源:本作作者] (Impact of flight on evolution:演化出的飞行能力的影响;Flight:飞行;Metabolism:新陈代谢;Reactive oxygen species:活性氧的种类;Cell damages: DNA, lipids, proteins:细胞损伤:DNA,脂质,蛋白质;Positive selection:正向选择;Reduction of inflammation in bats:蝙蝠炎症的减少;Mobilisation of interleukins to curb the interferon response:动员白细胞介素来抑制干扰素响应;decrease in the interferon regulation factor(IRF3):干扰素调节因子的降低;Decrease in the interferon beta:干扰素β的降低;Low level of inflammation:炎症等级降低;Advantageous for virus replication:利于病毒复制;Bat cell:蝙蝠细胞;Virus replication:病毒复制;Virus:病毒;Control of virus attack:抵御病毒攻击;Higher basal level for interferon expression in bats:蝙蝠干扰素更高基础等级的表达)
[1] Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, Si HR, Zhu Y, Li B, Huang CL, Chen HD, Chen J, Luo Y, Guo H, Jiang RD, Liu MQ, Chen Y, Shen XR, Wang X, Zheng XS, Zhao K, Chen QJ, Deng F, Liu LL, Yan B, Zhan FX, Wang YY, Xiao GF & Shi ZL. (2020). A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature, 579 (7798), 270-273. <doi: 10.1038/s41586-020-2012-7>.
[2] Xiao K, Zhai J, Feng Y, Zhou N, Zhang X, Zou J-J, Li N, Guo Y, Li X, Shen X, Zhang Z, Shu F, Huang W, Li Y, Zhang Z, Chen R-A, Wu Y-J, Peng S-M, Huang M, Xie W-J, Cai Q-H, Hou F-H, Chen W, Xiao L & Shen Y. (2020). Isolation of SARS-CoV-2-related coronavirus from Malayan pangolins. Nature, 583 (7815), 286-289. <doi: 10.1038/s41586-020-2313-x>.
[3] Calisher CH, Childs JE, Field HE, Holmes KV & Schountz T. (2006). Bats: important reservoir hosts of emerging viruses. Clin. Microbiol. Rev., 19 (3), 531-545. <doi:10.1128/CMR.00017-06>.
[4] Middleton DJ, Morrissy CJ, van der Heide BM, Russell GM, Braun MA, Westbury HA, Halpin K & Daniels PW. (2007). Experimental Nipah virus infection in pteropid bats (Pteropus poliocephalus). J. Comp. Pathol., 136 (4), 266-272. <doi: 10.1016/j.jcpa.2007.03.002>.
[5] Hu D, Zhu C, Wang Y, Ai L, Yang L, Ye F, Ding C, Chen J, He B, Zhu J, Qian H, Xu W, Feng Y, Tan W & Wang C. (2017). Virome analysis for identification of novel mammalian viruses in bats from Southeast China. Sci. Rep. 7 (1), 10917. <doi: 10.1038/s41598-017-11384-w>.
[6] Drexler JF, Corman VM, Müller MA, Maganga GD, Vallo P, Binger T, Gloza-Rausch F, Rasche A, Yordanov S, Seebens A, Oppong S, Adu Sarkodie Y, Pongombo C, Lukashev AN, Schmidt-Chanasit J, Stöcker A, Carneiro AJ, Erbar S, Maisner A, Fronhoffs F, Buettner R, Kalko EK, Kruppa T, Franke CR, Kallies R, Yandoko ER, Herrler G, Reusken C, Hassanin A, Krüger DH, Matthee S, Ulrich RG, Leroy EM & Drosten C. (2012). Bats host major mammalian paramyxoviruses. Nat. Commun., 3, 796. <Law: 10.1038/ncomms1796>.
[7] Sulkin SE & Allen R. (1974). Virology. Vol. 8, Basel: Melnick JL, Karger.
[8] Luis AD, Hayman DT, O’Shea TJ, Cryan PM, Gilbert AT, Pulliam JR, Mills JN, Timonin ME, Willis CK, Cunningham AA, Fooks AR, Rupprecht CE, Wood JL & Webb CT. (2013). A comparison of bats and rodents as reservoirs of zoonotic viruses: are bats special? Proc. Biol. Sci. 280 (1756), 20122753. <Law: 10.1098/rspb.2012.2753>.
[9] Olival KJ, Hosseini PR, Zambrana-Torrelio C, Ross N, Bogich TL & Daszak P. (2017). Host and viral traits predict zoonotic spillover from mammals. Nature, 546 (7660), 646-650. <doi: 10.1038/nature22975>.
[10] Anthony SJ, Johnson CK, Greig DJ, Kramer S, Che X, Wells H, Hicks AL, Joly DO, Wolfe ND, Daszak P, Karesh W, Lipkin WI, Morse SS; PREDICT Consortium, Mazet JAK & Goldstein T. (2017), Global patterns in coronavirus diversity. Virus Evol. 3 (1), vex012. <Law: 10.1093/ve/vex012>.
[12] Mollentze N & Streicker DG. (2020). Viral Zoonotic Risk Is Homogenous Among Taxonomic Orders of Mammalian and Avian Reservoir Hosts. Proc. Natl. Acad. Sci. USA, 117 (17), 9423-9430. <doi: 10.1073/pnas.1919176117>.
[13] Jebb D, Huang Z, Pippel M, Hughes GM, Lavrichenko K, Devanna P, Winkler S, Jermiin LS, Skirmuntt EC, Katzourakis A, Burkitt-Gray L, Ray DA, Sullivan KAM, Roscito JG, Kirilenko BG, Dávalos LM, Corthals AP, Power ML, Jones G, Ransome RD, Dechmann DKN, Locatelli AG, Puechmaille SJ, Fedrigo O, Jarvis ED, Hiller M , Vernes SC, Myers EW & Teeling EC. (2020). Six reference-quality genomes reveal evolution of bat adaptations. Nature, 583, 579-584. <doi: 10.1038/s41586-020-2486-3>.
[14] Zhang T, Wu Q & Zhang Z. (2020). Probable Pangolin Origin of SARS-CoV 2 Associated with the COVID-19 Outbreak. Curr. Biol. 30 (7), 1346-1351.e2. <doi: 10.1016/j.cub.2020.03.022>.
[15] Damas J, Hughes GM, Keough KC, Painter CA, Persky NS, Corbo M, Hiller M, Koepfli KP, Pfenning AR, Zhao H, Genereux DP, Swofford R, Pollard KS, Ryder OA, Nweeia MT, Lindblad-Toh K, Teeling EC, Karlsson EK & Lewin HA. (2020). Broad host range of SARS-CoV-2 predicted by comparative and structural analysis of ACE2 in vertebrates. Proc. Natl. Acad. Sci U.S.A. 2020 Aug 21:202010146. <doi: 10.1073/pnas.2010146117>.
[17] Brook CE & Dobson AP. (2015). Bats as special reservoirs for emerging zoonotic pathogens. Trends Microbiol. 23 (3), 172-180. <doi: 10.1016/j.tim.2014.12.004>.
[18] Zhang G, Cowled C, Shi Z, Huang Z, Bishop-Lilly KA, Fang X, Wynne JW, Xiong Z, Baker ML, Zhao W, Tachedjian M, Zhu Y, Zhou P, Jiang X, Ng J, Yang L, Wu L Xiao J, Feng Y, Chen Y, Sun X, Zhang Y, Marsh GA, Crameri G, Broder CC, Frey KG, Wang L-F & Wang J. (2013). Comparative analysis of bat genomes provides insight into the evolution of flight and immunity. Science, 339 (6118), 456-460. <doi: 10 .1126/science.1230835>.
[19] Banerjee A, Baker ML, Kulcsar K & Misra V, Plowright R, Mossman K. (2020). Novel insights into immune systems of bats. Front. Immunol. 11, 26. <doi: 10 .3389/fimmu.2020.00026>.
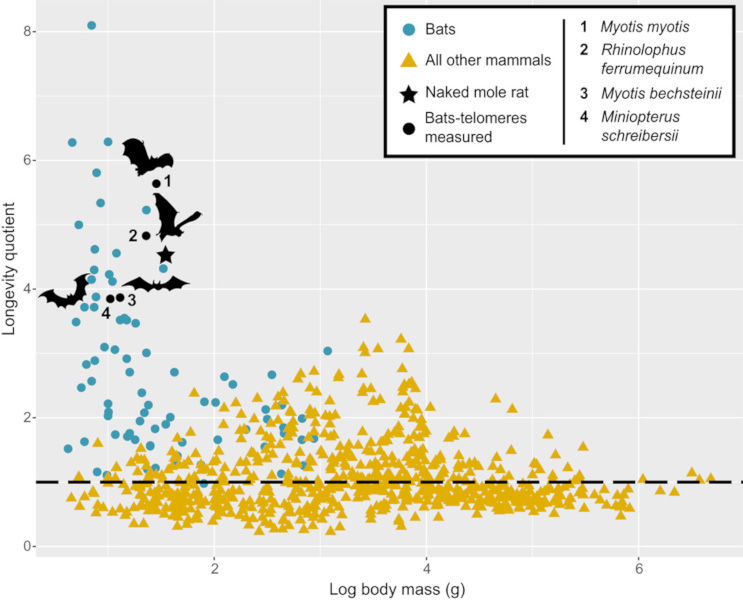
[20] Foley NM, Hughes GM, Huang Z, Clarke M, Jebb D, Whelan CV, Petit EJ, Touzalin F, Farcy O, Jones G, Ransome RD, Kacprzyk J, O’Connell MJ, Kerth G, Rebelo H, Rodrigues L, Puechmaille SJ & Teeling EC. (2018). Growing old, yet staying young: The role of telomeres in bats’ exceptional longevity. Sci. Adv. 4 (2), eaao0926. <10.1126/sciadv.aao0926>.
[21] Kacprzyk J, Hughes GM, Palsson-McDermott EM, Quinn SR, Puechmaille SJ, O’Neill LAJ & Teeling EC. (2017). A potent anti-inflammatory response in bat macrophages may be linked to extended longevity and viral tolerance. Acta Chiropter, 19 (2), 219-228. <doi: 10.3161/15081109ACC2017.19.2.001>.
[22] Mandle JN, Schneider C, Schneider DS, Baker ML. (2018). Going to bat(s) for studies of disease tolerance. Front Immunol, 9 (2112). < doi: 10.3389/fimmu.2018.02112>.
[23] Gibb R, Redding DW, Chin KQ, Donnelly CA, Blackburn TM, Newbold T & Jones KE. (2020). Zoonotic host diversity increases in human-dominated ecosystems. Nature, 584 (7821), 398-402. <doi: 10.1038/s41586-020-2562-8>.
[24] Afelt A, Frutos R & Devaux C. (2018). Bats, Coronaviruses, and Deforestation: Toward the Emergence of Novel Infectious Diseases? Front. Microbiol. 9, 702. <doi: 10.3389/fmicb.2018.00702>.