




第一代细胞的起源:工程师的观点
在生命起源的现实场景中,最初的新陈代谢始于水中矿物质的表面。它们促进了一些基本构建模块的聚合,包括RNA。这个由RNA代谢构成的世界逐渐发展,并导致这些RNA的复制,最终形成一个RNA 基因组世界,然后使用RNA分子作为信息模板(原始基因),而不是作为新陈代谢的直接底物。下一步的发展基于膜脂的出现,这是一种将内部介质与外部介质的分子。第一个细胞随之形成,将代谢和基因组分离的两个区室聚集在一起;分别是细胞质和细胞核。RNA分子的一种变体,即广为人知的DNA,促使基因组成染色体。膜脂促使的吞噬作用将成为真核细胞的起源,植物和动物都演化自真核细胞。
1.生命的起源:区分增殖和复制
许多以科学为基础的设想都提出了解决生命起源之谜的方案。但事实上,它们只是反映了作者的观点(参见《一位天文爱好者兼地质学家眼中的生命起源》和《生命出现的时候:40亿年前地球海洋中的化学成分》)。目前的情况也不例外。在进一步讨论之前,我们必须排除一个典型的陷阱,并决定我们是否将设想局限于地球,还是在宇宙的其他地方寻找生命起源。常见的解决方法是将地球生命的来源归结于外星生命。然而,这是一种通过转移问题来回避问题的方式,使问题更加受制于纯粹的幻想。因此,我们将遵循奥卡姆剃刀定律[1],根据我们对地球大气层和过去水环境的了解,来观察生命是否可能存在。我们还假设,生物体并不是“再生羊皮纸卷”(将已经使用的羊皮纸卷上的字迹刮去再次使用并写成手稿),被抹去了所有过去的痕迹,而是直到今天仍然包含着过去的档案。此外,我们将把我们的探索限制在生命的基本单元,即细胞,而不考虑多细胞生物。
在《生命的起源》这本鲜为人知的书中,物理学家弗里曼·戴森(Freeman Dyson)[2]指出,与生命相关的信息沿着两个截然不同的维度传播:
1.新陈代谢的增殖(产生相似的副本)(在一个有机体、器官或细胞内发生的所有生化反应,使有机体能够维持自身的生命、增殖、发展并对其环境做出反应。)(碳基分子的化学转化过程)。
2.程序的复制(至少在相当长的一段时间内产生相同的副本)。
更妙的是,他还进一步论证了在生命起源的真实场景中,原始过程的增殖必须先于复制过程的出现,然后它们联系成一个连贯的整体。在像地球表面这样的化学世界中,这意味着化学流的增殖和从这种新陈代谢中出现的能够自我复制的实体。简而言之,至少需要两个不同的化学起源来解释生命的起源。
2.利用基因组研究第一代细胞的起源
这种功能性的设想是非常抽象的,我们需要把它具体到地球的物理化学现实中。对细胞的分析表明,无论细胞如何起源,它总是由两类分子组成,这些分子都是由有限数量的原子(碳、氮、氧、氢、磷和硫)组成的。图 1列出了地球上现有的化学元素中的一些,其中少数发现于生命分子中。
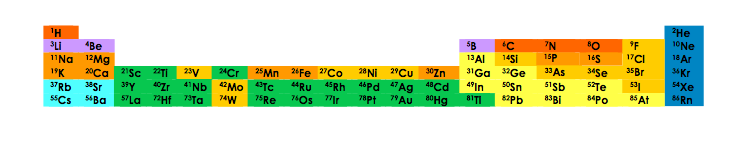
用深橙色表示的是组成蛋白质骨架的四种原子。橙色表示生命所必需的其他原子。浅橙色表示存在于许多生物体中的原子。铁元素几乎无处不在(虽然某些细菌种类中不含铁)。地球上大多数其他元素在生命分子中并不发挥关键作用。
分子可能由几个原子组成(“小”分子,代谢物),也可能是由数百万,甚至数十亿原子组成的大分子。这里有两类分子例外,一类是蛋白质,由20种基本单位(氨基酸)组成的序列,另一类是核酸,由4种不同单位(核苷酸)的链所组成。如果将大分子序列解读为一个用字母体系书写的文本,那么蛋白质由20个字母组成,而核酸由4个字母组成。因此,目前的中间代谢(负责小分子的合成)产生了期刊语言中常见的词汇——“砌块”:蛋白质中的氨基酸,以及参与核酸合成的核苷酸:核糖核酸(RNA),是由一连串的核糖核苷酸(腺嘌呤、胞嘧啶、鸟嘌呤、尿嘧啶)组成的大分子,在细胞内发挥许多功能。脱氧核糖核酸(DNA),是一种含有生物体遗传信息的大分子。由两条相互缠绕的反平行链组成,其形成双螺旋结构。其由核苷酸单体组成,由一个含氮碱基(腺嘌呤、胞嘧啶、鸟嘌呤或胸腺嘧啶)与脱氧核糖相连,脱氧核糖本身与磷酸基团相连。核苷酸变体在此不作讨论。为此,它使用了几种辅酶分子作为某些反应的辅助因子,这些反应是由酶催化的,它们在结构上与酶连接在一个稳定的复合物中,对于催化元素加速或减缓化学反应的作用至关重要。最后,小分子的世界必然先于大分子的世界出现,这是常识。
因此,任何关于生命起源的设想都应该解释这些代谢物的出现。然而,这些代谢物中的大多数在绝大多数的设想中都没有出现:尽管碳化学在宇宙中很常见,但许多作者对各种环境(通常离地球很远)中出现的几个氨基酸(并非全部氨基酸,实际上只有寥寥数个)感到欣喜若狂。奇怪的是,没有人调查它们是否伴随着相关的分子,或者与生命无关的分子(因此必须像毒药一样放在一旁),或者二者是否都存在。而对于辅酶的起源,以及很长一段时间内对于核苷酸起源,没有人提出质疑。最后,脂质是疏水性或两亲性分子(有一个疏水部分和一个亲水部分),其特点是不溶于水,可溶于非极性有机溶剂。这一对水显示出新特性的分子(一端亲水, 另一端疏水)用来形成细胞的被膜。人类几乎从未深入思考过这些分子的起源。然而,我们必须想到,从有限的基本成分(包括辅酶和脂质)中,原始的新陈代谢发展并增殖为中间代谢的雏形,直到代谢产物(可能是聚合物,蛋白质和核酸的雏形)发现了在脂质膜包围的空间框架内复制的方法。目前的基因组,自生命起源以来一直是生命记忆的载体,它为我们提供了必要的思路,使我们能够理解在这些早期阶段可能发生了什么。
越来越多的基因组序列不断积累。像商博良(Champollion)用罗塞塔石碑来理解象形文字那样将它们相互比较,使我们能够辨识现存基因组中共同的片段,也很有可能是古老的片段。不幸的是,进化倾向于保留功能,而不是结构,这使得这项工作更加困难。多个事物可能起到相同的作用(例如我们用叉子或筷子都可以吃饭)。另外,随着新基因组的测序,被认为对生命至关重要的基因数量不断减少。现在已经缩减到没有了!幸运的是,比较法并非完全失败,因为有些基因即使不存在于全部基因组中,也往往在许多基因组中保持存在。这些“坚持不懈”的基因编码了保守蛋白,因为它们有效地执行了最核心的功能。我们能对此说些什么?在细菌基因组中,基于计算机的观察表明,它们可以按照三个相互吸引的网络组合在一起。
核心网络是一个高度联系的网络,将管理其程序中遗传信息表达的基因聚集成簇。例如,我们在那里发现了驱动核糖体组装的基因,即携带该信息的 RNA 的通用阅读头。第二个联系较少的网络(起源较早)侧重于 RNA 的代谢。它定义了如今将信使 RNA(mRNA)翻译成蛋白质的酶,通过在同源的 RNA(转运RNA,tRNA)上装载二十种氨基酸中的每一种来实现。此外,联系其中的某些基因,我们发现了组织细胞分裂的基因。最后,第三组相互之间联系不紧密的基因编码了促使合成细胞中心代谢物、脂类、核苷酸和氨基酸的过程,还有酶蛋白的催化核心——辅酶。还有一些蛋白质的功能需要铁硫中心的存在,类似于一种常见矿物——黄铁矿的基本成分。最后,促使细胞膜脂质双分子层合成的基因也能在第三个网络中找到。
3.从矿物质到RNA
这种组织结构使人想到生命起源的现实情况,其基于对区室化的迫切需要,而不涉及生命之前的“汤”的任何情景。
3.1.石头的曙光:表面矿物质的选择和聚集
第一步,矿物质表面选择出活性化合物形成初级代谢。现存的代谢充分证明了这一观点:大多数中心代谢物是由带负电荷的分子(磷酸盐和羧酸盐)组成的,而很多时候,负责其电荷的化学基团在其功能中没有任何作用!因此,我们可以很自然地认为,这些基团显示出中心代谢物此前所起的作用,即分选和局部聚集矿物表面的分子,使它们能够相互反应。更重要的是,通过清除水分子,表面代谢为大分子的组装创造了驱动力,正如在肽(制造蛋白质)和磷酸二酯(制造核糖核酸)键的形成中观察到的那样。根据热力学第二原理,这种聚合的趋势源于环境中的水逃逸引起的熵增。水出于同样的原因(与普遍性的误解相反,是通过熵增,而不是熵减),在促进了大分子形成中作出了重要贡献。在这一步骤中,出现了氨基酸和催化中心,使得更复杂的代谢物得以形成。这个关于矿物质的设想可能围绕铁和硫(地球表面常见组织,最重要的分子是辅酶(对于含有这些辅酶的酶所加速的化学反应,这些辅酶是必要的催化物)、脂类(构建细胞膜必需的组分)和某些氨基酸。脂质的合成在水中自发地导致(同样得益于熵增)形成含有矿物质纳米颗粒的膜,构成原始的囊泡。
这些囊泡不断地融合以及分裂,既允许某些代谢物在局部聚集,又允许探索和共享各种代谢通路。当时的地球大气在电子转移方面是中性的。这意味着铁离子可溶于水(由于氧气的存在,今天已不再是这种情况),但同时又提出了基于碳骨架的分子中存在氮的问题。这一要求需要尽早出现固定大气中氮的机制,而氮气一种非常不活跃的气体。该阶段也是磷酸盐与多磷酸盐——这些矿物质能量非常丰富,但在水中呈亚稳态,承担着储存和化学能量转移的作用,使得代谢的动态组织成为可能。该阶段也见证了富含氮和磷的核苷酸的起源。它们目前的代谢与有机化学家在实验室合成这些分子时的代谢完全不同。它涉及产生肽的氨基酸作为前体。在遗传信息的翻译之外,是否还同时存在合成肽的反应?当然有,而且它们是围绕着涉及硫的反应产生的,这正是生物化学家克里斯汀·德迪夫(Christian de Duve)提出的生命的起源理论[3]。
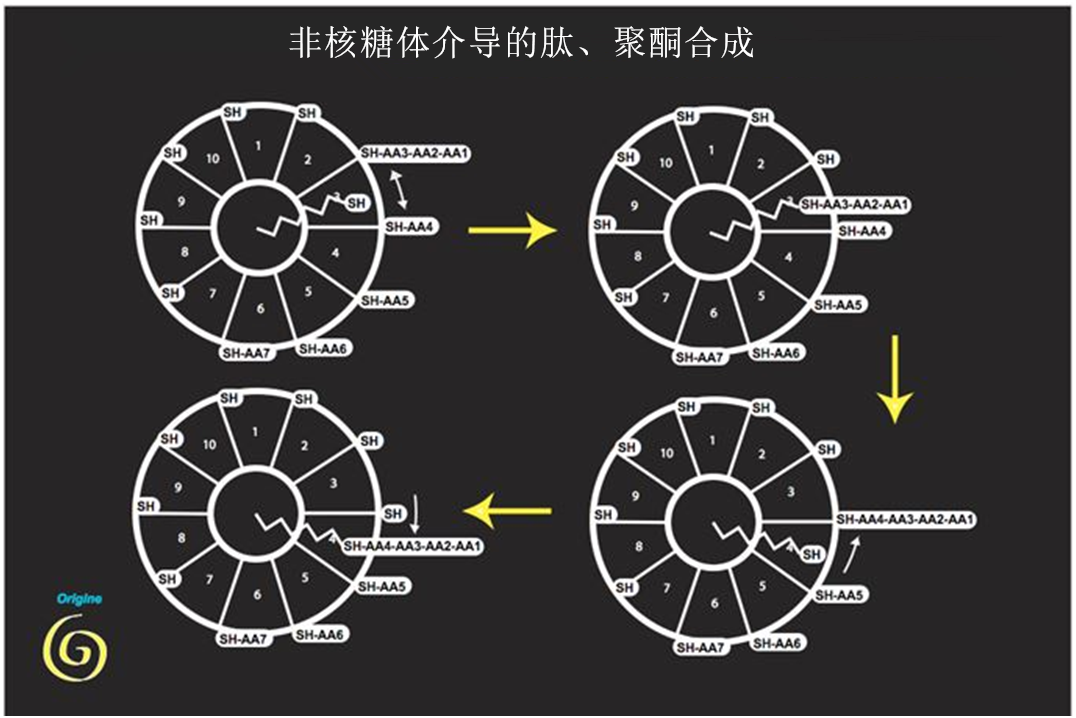
固定在支架上(最初是矿物质)上带有活性硫原子的柔性臂将底物从一个催化中心移到另一个催化中心,进行基本前体的局部聚合。直到今天肽、脂肪酸和更复杂的分子仍是这样构建的,这些分子参与辅酶和核苷酸的合成,更多和更复杂的合成由此产生。
这些反应产生了各种结构的肽(通常是抗生素)。这些肽有几个古老的特征:除了使用碳硫键这一关键特征外,它们还使用氨基酸的两种立体异构体(我们知道生命在蛋白质中只使用左手的异构体)。但有一项观察结果使这一假设更加引人注目:对合成脂肪酸(脂质的主要成分)的基因序列的分析表明,属于同一家族的酶(和相同的辅酶)促使它们的合成,证明了它们的共同起源(图 2)。在这里,我们首次解释了作为肽合成过程中意外副产品的膜的产生(其对区室化至关重要)。
3.2.“RNA代谢”的世界
第二个阶段建立在这种代谢的基础上,即“RNA 代谢”(具有催化各种代谢物之间反应能力的 RNA 分子,即核酶。)的世界。由核糖核苷酸聚合形成的RNA逐渐替代表面,成为刚性支持,使各种底物在局部发生变化。转运 RNA 的祖先家族,是信使 RNA 和核糖体内的蛋白质之间的适配器,代表了这些替代物中最合理的一类。实际上,这些分子的骨架今天被各种代谢物大规模修改,有许多例子表明,tRNA 以与蛋白质合成无关的方式介入反应,这正是它过去功能的现实体现。与此同时,成为核糖酶的RNA分子逐渐发现,与蛋白酶一样,它们可以催化各种代谢物之间的反应。在这种情况下,肽的形成,最初是随机的,似乎是RNA分子成为核糖体的关键反应。核糖体核酶的祖先最初在形成肽键的过程中将tRNA 祖先作为氨基酸保持装置。随后,当 RNA 基质对装载氨基酸的 tRNA 施加严格的更迭顺序时,肽中氨基酸序列的特异性得以显现。最后,RNA 自身的三维折叠,对于核酶的产生是必要的,这导致了序列互补性规律的发现(形成互补的双螺旋,其中核苷酸是互补的)。然后,这种将特定的 tRNA 与 RNA 模板的序列联系起来的互补规律,逐渐具体化为核苷酸三联体和氨基酸之间的刚性对应形式,导致了氨基酸-核苷酸密码的产生以及形成遗传密码的规则。
3.3.复制的发明:“RNA基因组”的世界
含有编码 RNA 分子的囊泡反复分裂和融合,与相关的肽编码组装系统同时复制和传播越来越高效的代谢途径。这场革命改变了事物的走向:基于 RNA 作为(可复制的)信息模板的“RNA 基因组”(RNA 以双螺旋的形式复制。根据互补定律,RNA 的副本作为生成多肽的基质)世界,从 RNA 代谢世界中不可逆转地分离出来。根据互补定律,可形成RNA模板的副本,呈现为RNA双螺旋的形式,用于肽的生成。肽的出现促进了它的复制,打开了 RNA 基因组的世界,其中 RNA 的转录和双螺旋的复制(获得与原始分子相同的两个分子的过程)仍然是重叠的过程。转录过程产生了原核糖体解码信息的模板,而复制则增加了这种基因的RNA祖先的副本数量。随后形成了含有各种成套序列的 RNA 双螺旋的囊泡,正如目前在一些 RNA 病毒的基因组中发现的那样。然而,这些原始细胞依赖于核糖核苷酸和多核糖核苷酸的持续合成,这些分子的化学性质非常不稳定。这个连续的过程要求在同一个区室(原始细胞)内既要有 RNA 代谢世界的复制,又要有 RNA 基因组世界的具体复制。因此,RNA 基因组世界必须找到一种方法来稳定其前体的合成,以避免消失。比核糖更稳定的脱氧核糖的出现解决了这个难题。因此,DNA 是随着时间推移非常稳定的分子,在记忆促进代谢产生方法的过程中,直到最后阶段才出现。这种新的分子将记忆性与细胞的一般功能分开。随后,编码 RNA 的 DNA 副本融合在一起,形成了第一条染色体。
4.第一代细胞
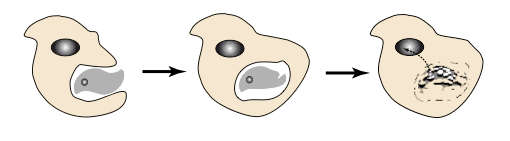
这些细胞由细胞质(来自 RNA 代谢)和细胞核(始于 RNA 基因组,后来被 DNA 取代)组成,吞噬同类型的其他细胞。这使它们能够捕获对其新陈代谢至关重要的元素,同时也能恢复含有在其他地方出现的编码的DNA 片段。因此,“水平基因转移”是该阶段进化的规则。
最后这一阶段见证了第一代细胞的诞生。为了理解它们的出现,有必要探索被脂质双分子层包围的囊泡是如何相互作用的。它们可以分裂和融合,但它们具有一个不变的特性,即相互渗透的能力,正如目前在吞噬作用(允许细胞包围并随后消化外来物质或生物体(如细菌)的过程)中观察到的那样。这一过程极大地丰富了代谢系统的进化,因为它允许在一个细胞内将具有不同但互补功能的区室联系起来。第一代细胞的祖先是吞噬细胞,将RNA代谢祖先的具有遗传信息的翻译和中间代谢的细胞质(细胞内部环境。它由水相和富含蛋白质的相(胞质溶胶)组成,并含有细胞器(线粒体等)),同RNA 基因组祖先的核心基因组联系在一起,特别是在出现 DNA 、以及细胞的代谢分为复制和转录为 RNA 之后。
因此,在进化的这一节点上,出现了一组吞噬细胞,即原核细胞(protokaryotes),它们通过增殖和系统地摄取它们周围的物质而进化。它们是巨大的残食同类者(就像今天的原生生物),通过它们的吞噬和传播,促进了代谢的革新。
这种情况是不稳定的。事实上,它导致了一种特定的、可以抵抗吞噬的功能形成。如果具有灵活的新陈代谢的生物体找到可以进行这种抵抗的结构,它将摆脱原核生物的残食,开始一个新的进化系。

膜脂将长链脂肪酸与甘油磷酸酯等分子结合起来。真核生物和细菌使用的这种骨架的立体异构体与古细菌使用的不同。其结果是,虽然后者的膜能以有限的方式与前者的膜合并,但属于它们的蛋白质分子(转运蛋白、保护层等)却不能合并。
对这种屏障有两种解决方案:用一种很难摄取的被膜将细胞包围,或者通过物理化学手段使吞噬作用不可能发生。细菌是找到第一种解决方案的细胞的后代,形成小细胞并被一个抗性被膜所包围。古细菌则发现如何用一种不可能被摄取的膜作为功能结构来包围自己,即使用和它们天敌的脂质在三维结构上镜像的脂质。它们通过在极端环境中定居进一步逃逸(图 4)。
总而言之,稳定的脱氧核糖核苷酸的出现使染色体内的基因得以分组,而基于古细菌(单细胞的原核微生物,特别是生活在极端环境中(厌氧、高盐、高热等)。卡尔·沃斯(Carl Woese)和乔治·E·福克斯(George E. Fox)(1977)的系统学研究区分了古细菌和其他原核生物(细菌)。目前,生物体被认为由三类组成:古细菌、细菌和真核生物。)中膜脂结构的代谢替代物和它们克服极端环境的能力,以及细菌出现坚固和带有抗性的被膜,吞噬作用引发了逃逸现象。然后,随着细菌和古细菌使细胞小型化,一些细菌对共享代谢产生兴趣,简化了它们的被膜,使它们中的一些成为原核生物的共生体(让人想起一些细菌在植物根部形成结节的方式)。对还原进化的追求将这种共生关系转变为将细菌基因组减少到线粒体(真核生物特有的细胞器)中的一小部分基因。单细胞或多细胞生物,其细胞拥有细胞核和由膜划界的细胞器(内质网、高尔基体、各种质体、线粒体等)。真核生物与细菌和古细菌一样,是三类生物之一。(见《共生和进化》)。
请注意,这种设想可能会在实验中得到验证。如果在某处仍有原始生物的直接后代,就会出现这种情况。这种假设生物体的特点是具有一个细胞质和一个细胞核,但没有线粒体(也没有源自细菌的细胞器的痕迹)。由于现今线粒体的主要功能是形成许多酶活动所必需的铁硫簇,因此必须在容易获得这些结构的环境中寻找这些生物。如果有这样一个结构存在,它将具有相当大的价值,可以将目前的设想从有根据的猜测转变为科学事实。
5.关于生命起源的设想
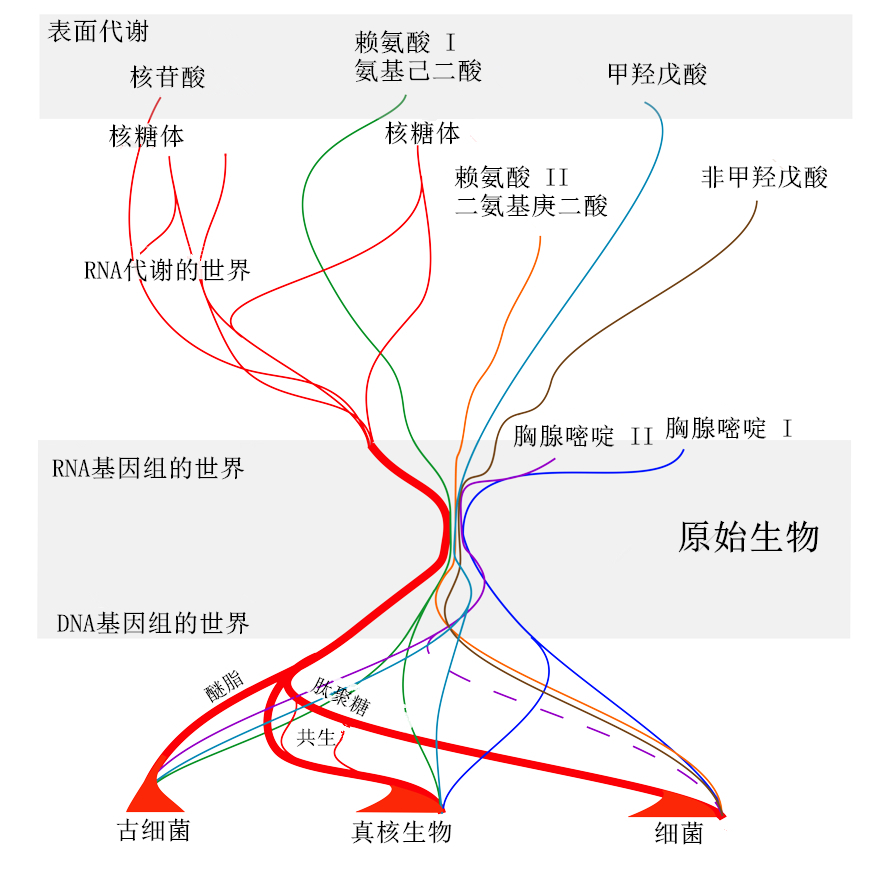
在表面代谢步骤之后,最终出现了主要的代谢物(确切地说,除了氨基酸、核苷酸、辅酶和脂质之外),两个 RNA 世界(RNA 代谢世界和 RNA 基因组世界)共同创造了核糖体的祖先,将这两个世界耦合起来。几条合成途径产生了蛋白质氨基酸——赖氨酸、DNA 碱基胸腺嘧啶以及产生重要辅酶和脂质的异戊二烯衍生物。在发现 DNA 后,基因被组合成染色体,第一代细胞,即原始生物(protokaryotes),从它们的吞噬行为中进化出来。一种特殊的赖氨酸合成途径的出现(使用二氨基庚二酸作为前体,而不是氨基己二酸)使其中一些细胞形成了抗吞噬的包膜。同样,用与以前不同的甘油磷酸酯异构体合成磷脂,使古细菌的祖先能够逃避吞噬作用。目前的真核生物是在细菌被摄取但未被杀死,与原核生物宿主共生在一起后出现的。
总之,本文提出的起源设想(图 5)假设同一物种的个体都是彼此不同的。然而,它也假设是一个共同的程序决定了它们的构造。这个程序代代相传,不加修改(它可以复制),而单个细胞只进行增殖(它们彼此相似,而不是相同)。增殖和复制之间的这种区别对于理解生命是什么至关重要,即不可避免地产生变体和尽可能保持程序不变性之间的对话。
在第一代细胞起源时,一个化学反应流,一个可复制的原始的新陈代谢,产生了各种有希望的意外,但也有太多的错误,因而无法保持持续。后来,通过编码实现了功能相关结构的精确复制。新陈代谢在水中矿物质的表面开始。与原始“汤”中发生的情况不同,矿物质表面不仅保留了以碳原子为中心的化学创造中的极小部分,而且由于涉及水的熵驱动过程,还促进了其中一些砌块的聚合。
一些砌块产生了大分子,即 RNA,替代了矿物质的表面。一个 RNA 代谢的世界就这样发展起来。然后,它促进了这些 RNA 的复制,形成了 RNA 基因组的世界。后者将 RNA 作为信息模板(原始基因),而不是作为代谢的直接底物。
由于产生了将内部环境与外部环境分开的分子——膜脂,第一代细胞(原始生物(protokaryotes))将两个区室聚集在一起,一个来自 RNA 代谢的世界(细胞质),另一个来自 RNA 基因组的世界(细胞核)。这些细胞是吞噬细胞,能够吞噬它们所遇到的一切,因此非常迅速地传播代谢革新。随后,RNA 的一个变体,即广为人知的 DNA,促使在染色体内对基因进行分组。
最后,还存在一种转折,吞噬作用使得逃逸吞噬成为可能,这种逃逸基于细胞被膜的改变。可能存在两种逃逸方式。膜脂是促使吞噬作用的关键,但它们是不对称的。用它们的镜像异构体取代它们可以减少或消除吞噬作用。膜对称性的这种变化引发了古细菌的起源。用一种非常有抵抗力的结构覆盖脂质是另一种解决方案,这引发了细菌的起源。然而,吞噬细胞仍然找到了一种方法来摄取一些细菌而不完全杀死它们,这引发了真核生物的起源,其中包括我们所知的植物和动物。
参考资料和说明
封面照片:海拉细胞 [来源:©美国国立卫生研究院(NIH),通过维基资源共享]
[1]奥卡姆剃刀原理:哲学推理的原则,也叫简单性原则或准绳性原则。
[2]弗里曼-戴森(1986),《生命的起源》,剑桥大学出版社。1999年第二版(ISBN 0521626684)。
[3]克里斯蒂安德迪夫(1917-2013),比利时生物化学家,因发现细胞中以前未知的细胞器——溶酶体而于1974年获得诺贝尔奖。这些细胞器在分解不同类型的物质方面具有重要功能,如细菌和细胞损伤的部分。
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: DANCHIN Antoine (2024年3月12日), 第一代细胞的起源:工程师的观点, 环境百科全书,咨询于 2026年7月23日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/origin-of-the-first-cells-engineers-point-of-view/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。


