



Guyane et nouvelles technologies pour l’écologie
PDFLa révolution technologique et méthodologique en écologie a conduit à des résultats nouveaux et spectaculaires. Rappelons que ce passage progressif de « l’écologie opinel, bouts de ficelle » à « l’écologie technologique » a été balisé par Legay et Barbault, en 1995 [1]. Ne sont cités ici que quelques avancées récentes postérieures à la publication de cet ouvrage.
1. Le Lidar
La technologie du Lidar, [2] d’abord utilisée par le spatial, a été adaptée pour la télédétection aérienne en vue d’obtenir des « modèles numériques » de terrain. En forêt tropicale (forêts denses et humides) elle a permis d’avoir une estimation de la hauteur des arbres, de cartographie au niveau du sol et à cette occasion de détecter des artéfacts archéologiques, dont la découverte est en train de révolutionner notre connaissance du passé de cet immense territoire qu’est l’Amazonie (voir figure 4 dans Amazonie : un vaste écosystème en évolution permanente). [3]
2. Dispositifs d’accès et d’observation de la canopée : du terrain à l’espace

Cet accès à la canopée est nécessaire, par exemple pour mesurer les échanges gazeux forêt-atmosphère, mesures réalisées pour évaluer les rôles de ces forêts dans la dynamique climatique [6]. Cependant, l’instrumentation du dispositif COPAS ne peut donner que des informations complémentaires à celle apportée par les « tours à flux » répondant à des critères internationaux du réseau Fluxnet. [7]

3. La metagénomique
Le séquençage haut-débit de l’ADN, les progrès de la bioinformatique et des méthodes biométriques ouvrent de grandes perspectives pour l’évaluation de la biodiversité au niveau local et aussi de reconstitution historique (Lire Code-barres ADN pour caractériser la biodiversité).
Rappelons que le métagénome est constitué par des morceaux d’ADN, molécules assez stables, provenant de divers organismes et stockés dans le milieu, notamment dans les sols. C’est ainsi que des analyses ont été réalisées à partir de sols de trois origines, dont un échantillon prélevé aux Nouragues (Tableau) [9].
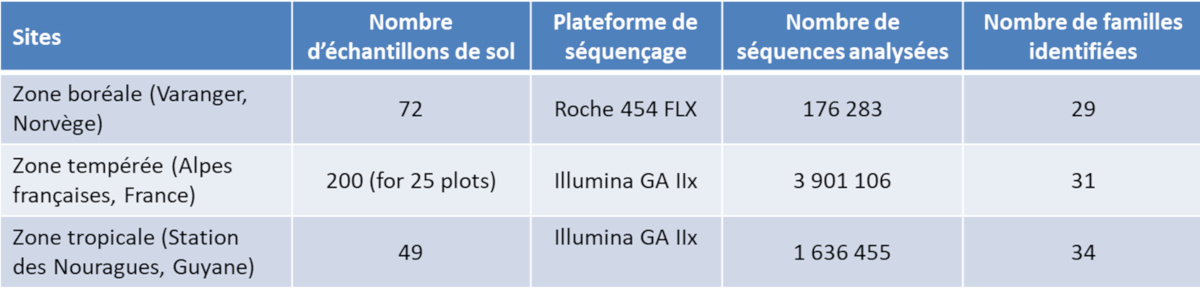
Tableau. Bilan d’analyses métagénomiques réalisées sur plusieurs sites, dont la Station des Nouragues en Guyane (voir réf. [9]).
L’utilisation des méthodes de séquençage permet d’établir une véritable collection de séquences permettant l’identification de l’espèce à laquelle appartient un organisme prélevé dans l’environnement. On envisage de fabriquer des kits d’identification moléculaire.
4. Les capteurs
Un effort important a été porté sur les capteurs pour l’écologie [10]. Les stations de terrain en sont équipés, par exemple aux Nouragues et à Paracou en Guyane : suivi d’animaux, mesures des échanges forêt-atmosphère en continu, stations météo automatiques, liaison internet par satellite, etc. [11] Ces équipements ne concernent pas uniquement les dispositifs de terrain, mais aussi les moyens aériens et spatiaux. C’est ainsi que la mission BIOMASS de l’ESA a été préparée en Guyane dans le cadre du projet TROPISAR [12], notamment pour calibrer le radar qui sera embarqué. La Guyane a été choisie car elle dispose, grâce aux stations de terrain gérées par les organismes de recherche, d’une qualité de suivi scientifique des massifs forestiers unique au monde. Le satellite assurant cette mission sera lancé de Kourou, en 2020, par le lanceur Vega.
L’image en en-tête de ce focus est une vue du site principal, dit « site inselberg », de la station des Nouragues en Guyane (100 km au sud de Cayenne). L’autre site à 5 km au sud, dit « site Arataye » est équipé, entre autre, du dispositif COPAS présenté ci-dessus. Les deux sites sont équipés de capteurs, notamment pour mesurer les paramètres atmosphériques (température, pression, humidité, composition en GES) et de liaisons satellite. On dispose aussi d’appareils photo à déclenchement automatique permettant de détecter des animaux et d’évaluer localement des nombres d’individus. C’est ainsi que l’ONSFS [13] a réévalué la population de jaguars à la hausse.
Enfin, les recherches menées en Guyane et plus généralement dans la région amazonienne concernent aussi les hydrosystèmes fluviaux et lacustres, ainsi que les littoraux. Dans ce dernier cas, on peut citer les travaux menés sur des animaux marins, notamment sur les tortues Luth qui sont mêmes utilisées comme porteuses de capteurs pour mesurer des paramètres marins (température, salinité, direction et vitesse du courant) au cours de leurs déplacement dans l’Atlantique nord.
5. Chimie des substances naturelles biologiquement actives
La chimie des substances naturelles biologiquement actives est essentielle, non seulement pour la recherche de ces substances avec des enjeux de valorisation, mais également leur rôle écologique, par exemple comme signaux comportementaux. Cette chimie met en jeu des techniques sophistiquées, comme la chromatographie en phase gazeuse couplée à la spectrométrie de masse (GC-MS), ainsi que la résonance magnétique nucléaire (RMN). Ces équipements ont été installés en Guyane et contribuent à l’identification de ces substances. Des coopérations existent avec des laboratoires brésiliens.
6. Modélisation et théorisation
Bien entendu, les modèles et théories de l’écologie ne sont pas propres à l’écologie des forêts tropicales, mais les objets d’études exhibent des propriétés amplifiées et spécifiques par rapport à leur équivalents tempérés. La première est due à leur faible anthropisation, les processus écologiques s’expriment de façon spontanée. Par exemple, la résilience de ces forêts est en grande partie la conséquence de leurs structures spatiales locale très largement aléatoire : le peuplement est très mélangé, très diversifié, de sorte qu’une perturbation locale n’oblitère pas la régénération forestière avec une diversité équivalente.
On pourrait citer bien d’autres exemples : l’écologie a résolument pris le virage technologique, méthodologique et théorique. Il reste néanmoins un long chemin à parcourir si l’on reprend la comparaison, néanmoins osée, avec l’astronomie. Cette dernière a l’avantage de disposer de références théoriques solides et mathématiquement formalisées, l’écologie en est encore loin. On peut néanmoins être surpris par les avancées avant la guerre, notamment en France, soulignées par Scudo et Ziegler [18]. Ensuite, il semble que le recours à la réflexion théorique ait été ralenti jusqu’aux années 1980, pour des raisons mal connues.
Notes et références
Image de couverture. [Source : © Alain Pavé]
1] Legay J.M. & Barbault R. (Dir.) (1995) La révolution technologique en écologie. Masson, Paris.
[2] Laser remote sensing or lidar, an acronym for the English term « laser detection and ranging », is a remote measurement technique based on the analysis of the properties of a beam of light reflected back to its transmitter. Unlike radar, which uses radio waves, or sonar, which uses acoustic waves, lidar uses light (visible spectrum, infrared or ultraviolet). It is almost always produced by a laser.
[3] Jérémie S. & Dambrine E. (2010) Impact des occupations amérindiennes anciennes sur les propriétés des sols et la diversité des forêts guyanaises. In, Alain Pavé & Gaëlle Fornet, Amazonie, une aventure scientifique et humaine du CNRS, Ed. Galaade, Paris.
[4] Projet réalisé en collaboration entre l’Université d’Ulm, la fondation Körber, l’UE et le CNRS ; voir http://www.guyane.cnrs.fr/IMG/pdf/FICHE_COPAS_JUILLET2014.pdf
[5] Le CNRS a fortement contribué au développement de certains instruments.
[6] Mesurer la forêt : http://www.guyane.cnrs.fr/IMG/pdf/Poster_AnAEE_EC_2016.pdf
[7] https://fluxnet.fluxdata.org/
[8] http://sites.fluxdata.org/GF-Guy/
[9] Taberlet P. et al (2012) Soil sampling and isolation of extracellular DNA from large amount of starting material suitable for metabarcoding studies. Mol. Ecol. 21:1816-1820; Yoccoz et al (2012) DNA from soil mirrors plant taxonomic and growth form diversity. Mol. School. 2012, 21, 3647-3655.
[10] Le Gaillard J.F., Guarini J.M. & Gaill F. (2012) Sensors for ecology. Towards integrated knowledge of ecosystems. CNRS-INEE, Paris.
[11] Chave J., Gaucher Ph. & Dewynter M. (2012) Tropical rain forest environmental sensors at the Nouragues experimental station in French Guyana. In Le Gaillard J.F., Guarini J.M., Gaill F. Sensors for ecology. Towards integrated knowledge of ecosystems. CNRS-INEE, Paris.
[12] http://www.cnes-csg.fr/automne_modules_files/csg_pub/articles/r517_Leader_Guyane_L5N86.pdf
[13] Office national de la chasse et de la faune sauvage
[14] Volkov I, Banavar J.R., Hubbell S.P. & Maritan A. (2003) Neutral theory and relative species abundance in ecology. Nature, 424, 1035-1037
[15] He F. & Hubbell S. P. (2011) Species-area relationships always overestimate extinction rates from habitat loss. Nature, 473, 368-371.
[16] Pavé A. (2019) Understanding biodiversity, real problems and misconceptions. Editions du Seuil, Paris.
[17] Pan X. (2013) Fundamental equations for species-area theory. Nature: Scientific reports, 1334, DOI:10:1038:srep01334.
[18] Scudo F.M. & Ziegler J.R. (1978) The Golden Age of Theoretical Ecology: 1923-1940. Read. Notes in Biomathematics, Springer-Verlag