


French Guiana and new technologies for ecology
PDFThe technological and methodological revolution in ecology has led to new and spectacular results. It should be recalled that this progressive transition from “opinel ecology, bits of string” to “technological ecology” was marked out by Legay and Barbault in 1995 [1]. Only a few recent advances since the publication of this book are mentioned here.
1. The Lidar
The Lidar technology [2], first used by space, has been adapted for aerial remote sensing to obtain “digital models” of the terrain. In tropical forests (dense and humid forests), it has made it possible to estimate the height of trees, map them at ground level and, on this occasion, detect archaeological artifacts, the discovery of which is revolutionizing our knowledge of the past of this immense territory that is the Amazon (see Figure 4 in Amazonia: a huge ecosystem in constant evolution) [3].
2. Canopy access and observation devices: from the ground to space

This access to the canopy is necessary, for example to measure the gas exchange between the forest and the atmosphere, measures taken to assess the roles of these forests in climate dynamics [5]. However, the instrumentation of the COPAS system can only provide additional information to that provided by “flow towers” meeting international criteria of the Fluxnet network [7].

3. Metagenomics
High-throughput DNA sequencing, advances in bioinformatics and biometric methods open up great opportunities for biodiversity assessment at the local level and also for historical reconstruction (see DNA Barcode to characterize biodiversity).
It should be remembered that the metagenome is made up of pieces of DNA, fairly stable molecules, from various organisms and stored in the environment, particularly in the soil. Thus, analyses were carried out on soils from three origins, including a sample taken in Les Nouragues (Table) [9].
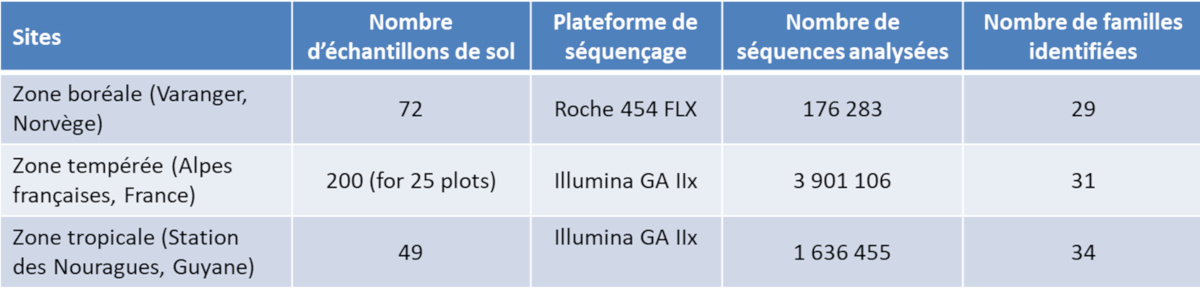
Table. Assessment of metagenomic analyses carried out on several sites, including the Nouragues Station in French Guiana (see ref. [9]).
The use of sequencing methods makes it possible to establish a real collection of sequences allowing the identification of the species to which an organism taken from the environment belongs. There are plans to manufacture molecular identification kits.
4. Sensors
A major effort has been made on sensors for ecology [10]. Field stations are equipped with them, for example in Nouragues and Paracou in French Guiana: animal monitoring, continuous forest-atmosphere exchange measurements, automatic weather stations, satellite internet connection, etc. [11]. This equipment does not only concern field devices, but also air and space assets. Thus, the ESA BIOMASS mission was prepared in French Guiana as part of the TROPISAR project [12]. in particular to calibrate the radar that will be on board. Guyana was chosen because it has, thanks to the field stations managed by research organisations, a quality of scientific monitoring of forest areas that is unique in the world. The satellite carrying out this mission will be launched from Kourou in 2020 by the Vega launcher.
The header image of this focus is a view of the main site, known as the “inselberg site”, of the Nouragues station in French Guiana (100 km south of Cayenne). The other site 5 km to the south, known as the “Arataye site”, is equipped with, among other things, the COPAS system presented above. Both sites are equipped with sensors, in particular to measure atmospheric parameters (temperature, pressure, humidity, GHG composition) and satellite links. There are also cameras with automatic triggering for detecting animals and locally assessing numbers of individuals. As a result, the ONSFS [13] has reassessed the jaguar population upward.
Finally, research carried out in French Guiana and more generally in the Amazon region also concerns river and lake hydrosystems, as well as coasts. In the latter case, we can mention the work carried out on marine animals, particularly on Luth turtles, which are even used as carriers of sensors to measure marine parameters (temperature, salinity, direction and speed of the current) during their movements in the North Atlantic.
5. Chemistry of biologically active natural substances
The chemistry of biologically active natural substances is essential, not only for the research of these substances with recovery issues, but also their ecological role, for example as behavioural signals. This chemistry involves sophisticated techniques such as gas chromatography coupled with mass spectrometry (GC-MS) and nuclear magnetic resonance (NMR). This equipment has been installed in French Guiana and contributes to the identification of these substances. Cooperation exists with Brazilian laboratories.
6. Modeling and theorization
Of course, ecological models and theories are not specific to tropical forest ecology, but the objects of study exhibit amplified and specific properties compared to their temperate counterparts. The first is due to their low anthropization, ecological processes are expressed spontaneously. For example, the resilience of these forests is mainly the result of their largely random local spatial structures: the stand is very mixed, very diversified, so that a local disturbance does not obliterate forest regeneration with equivalent diversity.
Many other examples could be cited: ecology has resolutely taken the technological, methodological and theoretical shift. Nevertheless, there is still a long way to go if we resume the comparison, nevertheless daring, with astronomy. The latter has the advantage of having solid theoretical and mathematically formalized references, ecology is still far from that. Nevertheless, we can be surprised by the progress made before the war, particularly in France, highlighted by Scudo and Ziegler [18]. Second, it seems that the use of theoretical reflection was slowed down until the 1980s, for reasons that are not well known.
Notes and references
Cover image. [Source: © Alain Pavé]
[1] Legay J.M. & Barbault R. (Dir.) (1995) La révolution technologique en écologie. Masson, Paris. (in French)
[2] Laser remote sensing or lidar, an acronym for the English term “laser detection and ranging”, is a remote measurement technique based on the analysis of the properties of a beam of light reflected back to its transmitter. Unlike radar, which uses radio waves, or sonar, which uses acoustic waves, lidar uses light (visible spectrum, infrared or ultraviolet). It is almost always produced by a laser.
[3] Jérémie S. & Dambrine E. (2010) Impact des occupations amérindiennes anciennes sur les propriétés des sols et la diversité des forêts guyanaises. In, Alain Pavé & Gaëlle Fornet, Amazonie, une aventure scientifique et humaine du CNRS, Ed. Galaade, Paris. (in French)
[4] Project carried out in collaboration between the University of Ulm, the Körber Foundation, the EU and the CNRS; see http://www.guyane.cnrs.fr/IMG/pdf/FICHE_COPAS_JUILLET2014.pdf
[5] The CNRS has made a significant contribution to the development of certain instruments.
[6] Measuring the forest: http://www.guyane.cnrs.fr/IMG/pdf/Poster_AnAEE_EC_2016.pdf
[7] https://fluxnet.fluxdata.org/
[8] http://sites.fluxdata.org/GF-Guy/
[9] Taberlet P. et al (2012) Soil sampling and isolation of extracellular DNA from large amount of starting material suitable for metabarcoding studies. Mol. Ecol. 21:1816-1820; Yoccoz et al (2012) DNA from soil mirrors plant taxonomic and growth form diversity. Mol. School. 2012, 21, 3647-3655.
[10] Le Gaillard J.F., Guarini J.M. & Gaill F. (2012) Sensors for ecology. Towards integrated knowledge of ecosystems. CNRS-INEE, Paris.
[11] Chave J., Gaucher Ph. & Dewynter M. (2012) Tropical rain forest environmental sensors at the Nouragues experimental station in French Guyana. In Le Gaillard J.F., Guarini J.M., Gaill F. Sensors for ecology. Towards integrated knowledge of ecosystems. CNRS-INEE, Paris.
[12] http://www.cnes-csg.fr/automne_modules_files/csg_pub/articles/r517_Leader_Guyane_L5N86.pdf
[13] Office national de la chasse et de la faune sauvage
[14] Volkov I, Banavar J.R., Hubbell S.P. & Maritan A. (2003) Neutral theory and relative species abundance in ecology. Nature, 424, 1035-1037
[15] He F. & Hubbell S. P. (2011) Species-area relationships always overestimate extinction rates from habitat loss. Nature, 473, 368-371.
[16] Pavé A. (2019) Understanding biodiversity, real problems and misconceptions. Editions du Seuil, Paris.
[17] Pan X. (2013) Fundamental equations for species-area theory. Nature: Scientific reports, 1334, DOI:10:1038:srep01334.
[18] Scudo F.M. & Ziegler J.R. (1978) The Golden Age of Theoretical Ecology: 1923-1940. Read. Notes in Biomathematics, Springer-Verlag