


La révolution des archées Asgard
PDF1. Des procaryotes distincts des bactéries
Jusqu’aux années 1970, les biologistes pensaient que tous les micro-organismes dépourvus de noyau appartenaient à un seul grand groupe : les bactéries. Cette vision fut bouleversée par une découverte qui modifia notre représentation de l’arbre du vivant et éclaira, plusieurs décennies plus tard, l’origine des cellules eucaryotes.
En 1977, le microbiologiste Carl Woese et son collègue George Fox ont réalisé une découverte majeure. En analysant l’ARN ribosomique 16S, une molécule présente chez tous les êtres vivants, ils ont montré que certains micro-organismes, jusqu’alors classés parmi les bactéries, formaient en réalité un groupe totalement différent : les archées [1].
Cette découverte a bouleversé la biologie pour trois raisons essentielles.
Premièrement, elle a conduit à une révision complète de l’arbre du vivant : avant Woese, tous les procaryotes (organismes sans noyau) étaient regroupés sous le terme « bactéries ». On sait désormais que bactéries et archées constituent deux lignées aussi éloignées l’une de l’autre que les bactéries le sont des eucaryotes. Deuxièmement, elle a profondément modifié notre vision de l’évolution : la diversité du vivant est plus ancienne et plus profonde qu’on ne le pensait. Les archées ne sont pas des bactéries particulières : elles constituent une lignée aussi distincte des bactéries que les eucaryotes le sont de celles-ci. Enfin, elle a ouvert un nouveau champ en écologie microbienne. Les archées occupent pratiquement tous les milieux :
- dans les environnements extrêmes (sources hydrothermales brûlantes, lacs hypersalés, milieux très acides) ;
- mais aussi dans les océans, les sols, les sédiments ;
- et même dans des milieux anaérobies (dépourvus d’oxygène), riches en matière organique, notamment dans le tube digestif de nombreux animaux.
Qu’est-ce qui les distingue concrètement des bactéries ?
Les archées ne possèdent pas de peptidoglycane dans leur paroi cellulaire. Leurs lipides membranaires sont liés au glycérol par des liaisons éther, contrairement aux lipides des bactéries et des eucaryotes, qui utilisent des liaisons ester. Surtout, leurs machineries moléculaires — ARN polymérase, mécanisme de traduction des protéines et protéines histones-like — sont beaucoup plus proches de celles des eucaryotes (les cellules à noyau, comme les nôtres) que de celles des bactéries.
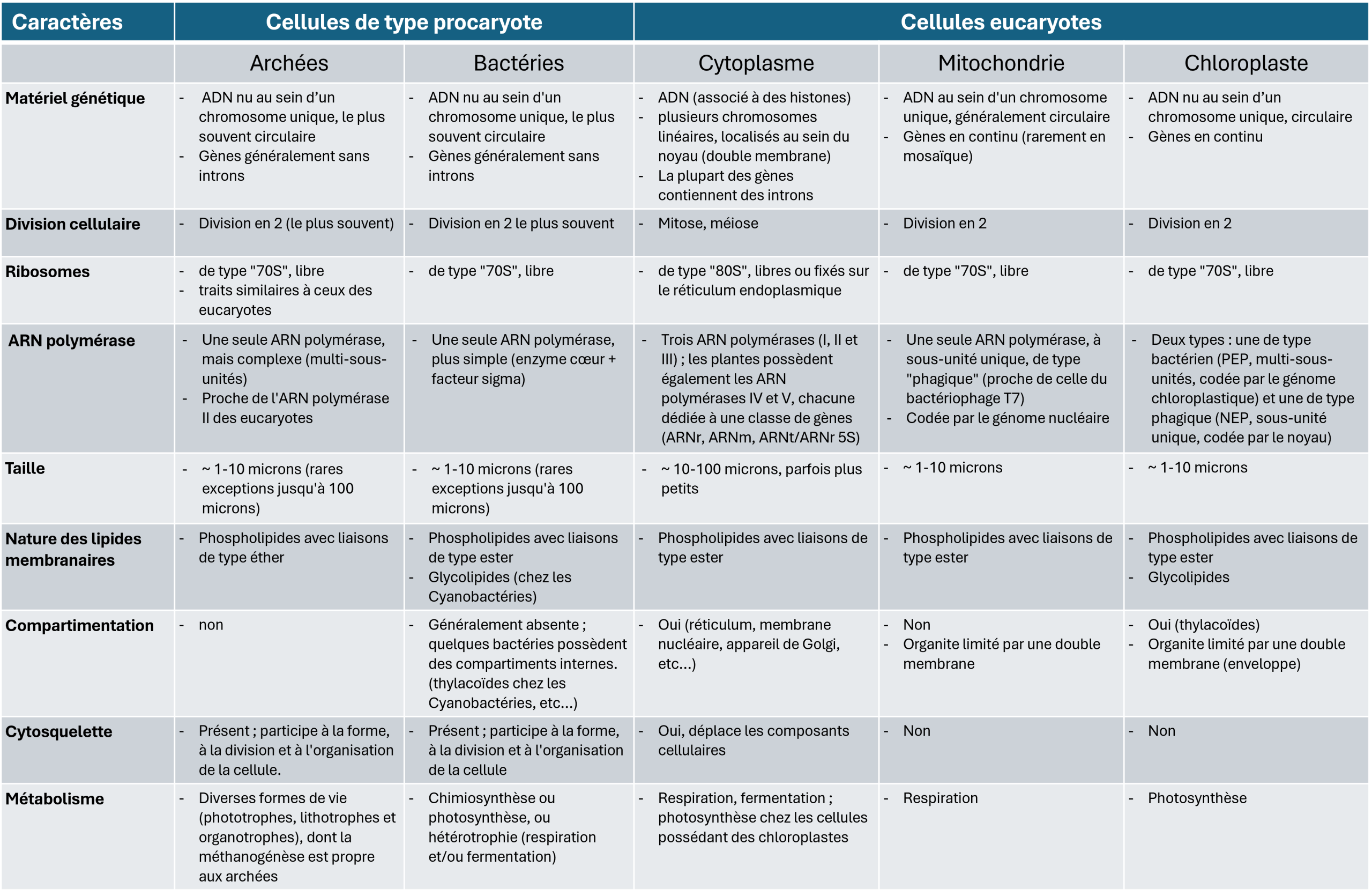
Tableau 1. Comparaison cellules de type eucaryote et procaryote (voir Symbiose et évolution : à l’origine de la cellule eucaryote).
Pendant près de quarante ans, les archées furent essentiellement considérées comme le troisième domaine du vivant. La découverte des archées Asgard allait profondément modifier cette vision.
2. Des gènes « eucaryotes » chez des microbes procaryotes
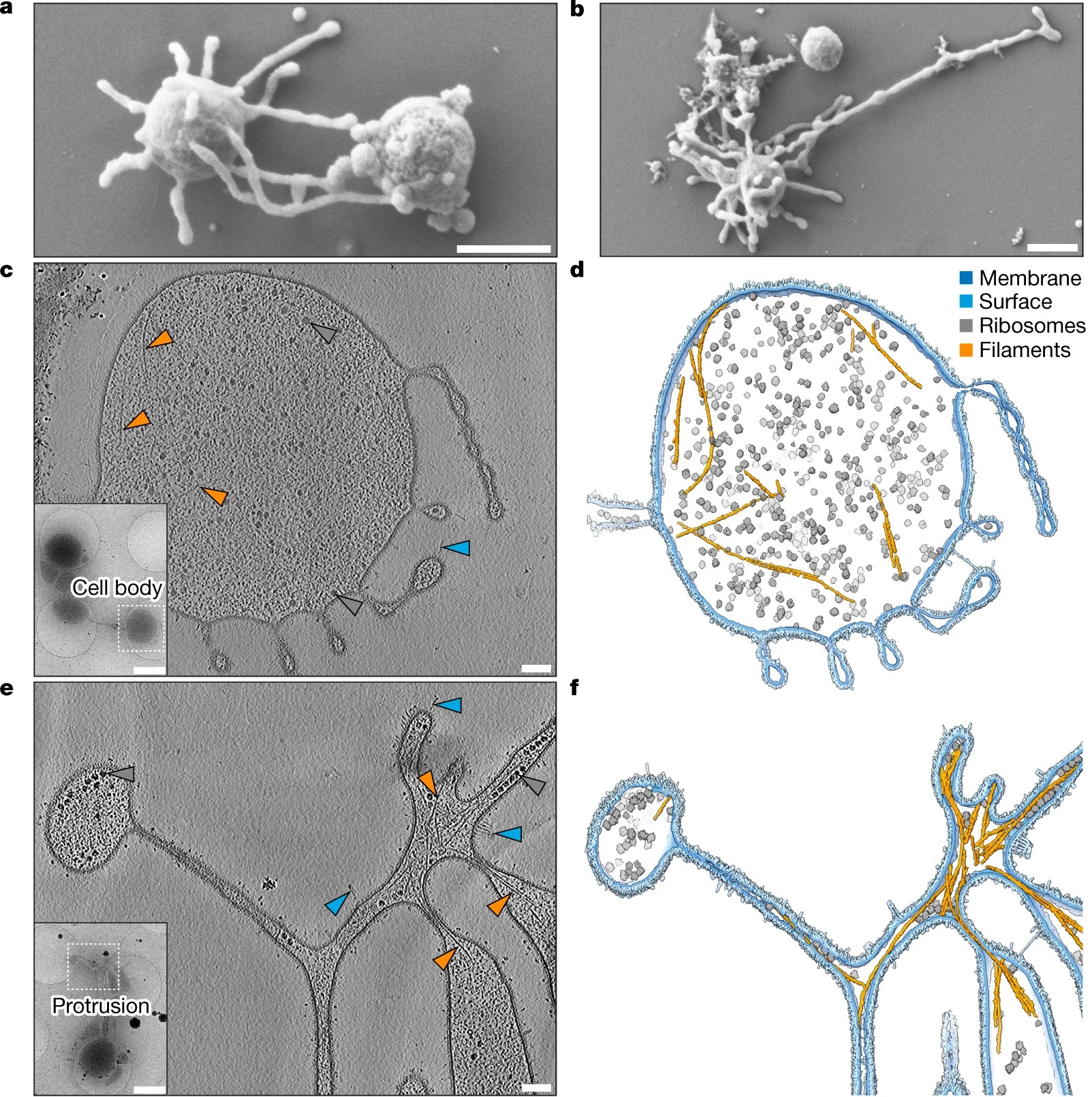
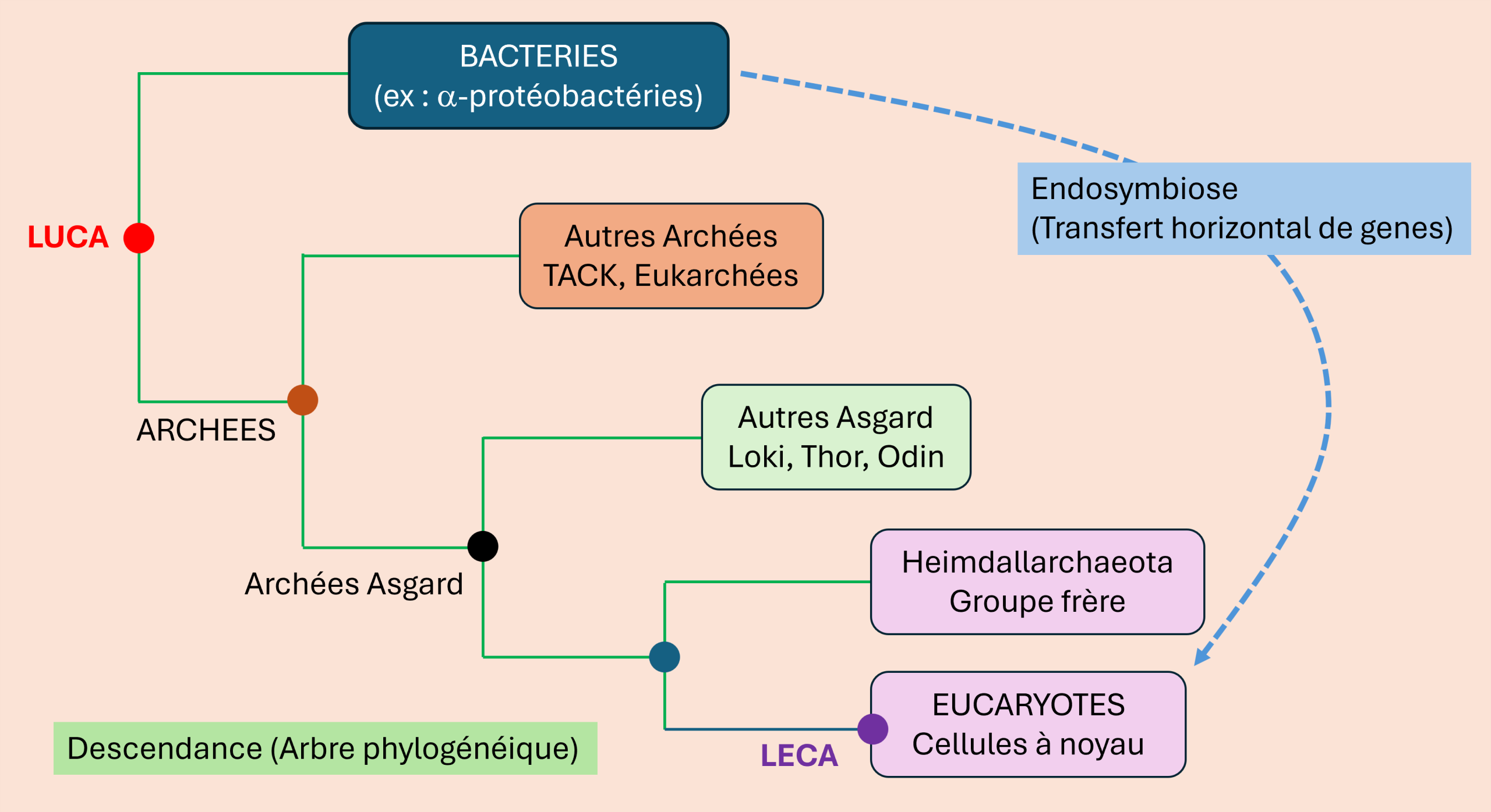
La grande révolution est venue en 2015. En analysant des sédiments prélevés près de l’évent hydrothermal Loki’s Castle (Figure 1) dans l’océan Arctique, des chercheurs ont reconstruit le génome d’un organisme jusqu’alors inconnu : Lokiarchaeota [2]. Deux ans plus tard, en 2017, une équipe suédoise a formalisé cette famille sous le nom d’archées Asgard — d’après le royaume des dieux nordiques — et dressé le premier catalogue de leurs gènes eucaryotes [3]. La découverte des archées Asgard a profondément renouvelé notre compréhension de l’origine des eucaryotes en révélant qu’un grand nombre de caractères moléculaires réputés propres aux eucaryotes existaient déjà chez leur ancêtre archéen.

D’autres lignées apparentées ont rapidement été identifiées (Heimdallarchaeota, Thorarchaeota, Odinarchaeota…). Ensemble, elles forment le super-groupe des archées Asgard. Grâce à la métagénomique (séquençage direct de l’ADN environnemental sans culture), les scientifiques ont pu explorer ces organismes impossibles à cultiver en laboratoire pendant longtemps.
Les analyses de centaines de génomes Asgard ont identifié plus de 500 protéines de signature eucaryote impliquées notamment dans le trafic vésiculaire, l’ubiquitine et le cytosquelette, montrant que plusieurs éléments de la complexité eucaryote existaient déjà chez leur ancêtre archéen.
3. Un nouveau modèle de l’origine des eucaryotes
Pourquoi cette transition a-t-elle été si exceptionnelle ?
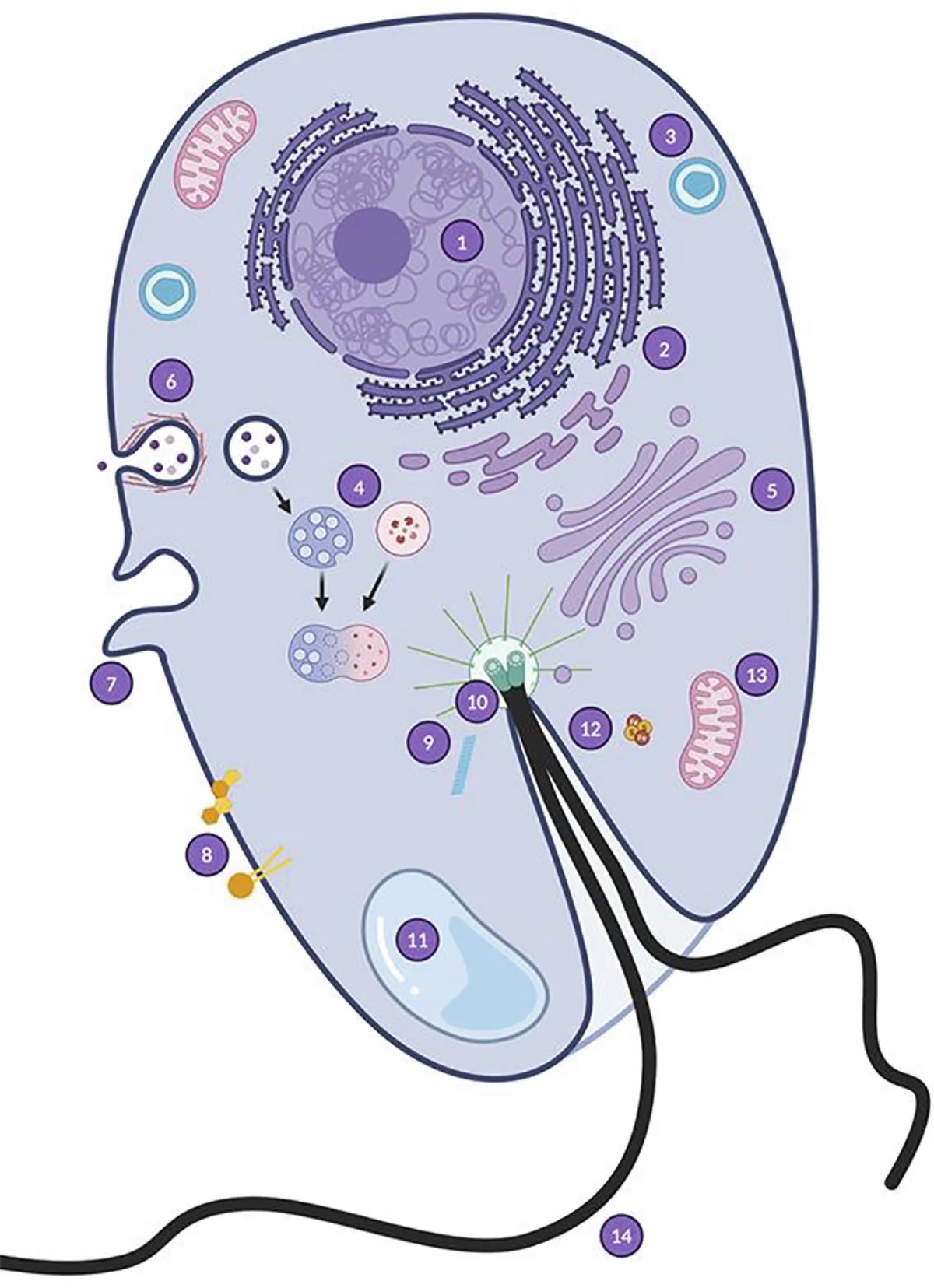
L’émergence des eucaryotes n’est arrivée qu’une seule fois dans l’histoire de la vie. Les Asgard montrent que l’hôte n’était pas une cellule simple et « passive », mais une cellule déjà pré-adaptée, dotée d’outils (cytosquelette, capacité à former des vésicules) qui ont permis d’intégrer durablement la mitochondrie. L’apport énergétique fourni par les mitochondries a ensuite permis de maintenir un génome plus grand, de produire plus de protéines et d’atteindre une complexité inédite.
De nombreuses questions restent ouvertes : le noyau s’est-il formé pour séparer transcription et traduction face à l’afflux massif de gènes bactériens ? Les virus ont-ils joué un rôle dans les transferts de gènes ou la formation du noyau ? La position exacte de la racine eucaryote au sein de l’arbre Asgard — proche des Heimdallarchaeota ou d’un groupe encore plus proche, les Wukongarchaeota — fait elle-même l’objet d’un débat actif [11]. Les futures cultures d’archées Asgard et analyses génomiques continueront d’affiner ce scénario.
Les archées Asgard ont profondément renouvelé notre compréhension de l’origine des eucaryotes. Elles montrent que la cellule eucaryote n’est pas apparue soudainement, mais résulte d’une longue histoire évolutive commencée au sein d’une lignée d’archées déjà remarquablement complexe, puis transformée par l’intégration durable d’une α-protéobactérie devenue mitochondrie. En quelques années, ces micro-organismes sont devenus des témoins privilégiés de l’une des plus grandes transitions de l’histoire de la vie. Leur découverte illustre de manière exemplaire la façon dont la métagénomique, la phylogénomique et la microbiologie expérimentale continuent de renouveler notre compréhension de l’évolution.
Notes & références
Vignette. Promethearchaeum syntrophicum aka Lokiarchaeota sp. MK-D1. SEM images of MK-D1 cells producing long branching membrane protrusions. Scale bars, 1 μm. [Source Imachi, H., Nobu, M.K., Nakahara, N. et al. Isolation of an archaeon at the prokaryote–eukaryote interface. Nature 577, 519–525 (2020). https://doi.org/10.1038/s41586-019-1916-6, licence CC BY 4.0 via Wikimedia Commons]
[1] Woese, C. R., & Fox, G. E. (1977). Phylogenetic structure of the prokaryotic domain: The primary kingdoms. Proceedings of the National Academy of Sciences, 74(11), 5088–5090.
[2] Spang A, Saw JH, Jørgensen SL, Zaremba-Niedzwiedzka K, Martijn J, Lind AE, van Eijk R, Schleper C, Guy L, Ettema TJG. Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature. 2015 May 14;521(7551):173-179. doi: 10.1038/nature14447.
[3] Zaremba-Niedzwiedzka, K. et al. Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature 541, 353–358 (2017). https://doi.org/10.1038/nature21031
[4] Imachi, H., Nobu, M.K., Nakahara, N. et al. Isolation of an archaeon at the prokaryote–eukaryote interface. Nature 577, 519–525 (2020). https://doi.org/10.1038/s41586-019-1916-6
[5] Rodrigues-Oliveira, T., Wollweber, F., Ponce-Toledo, R.I. et al. Actin cytoskeleton and complex cell architecture in an Asgard archaeon. Nature 613, 332–339 (2023). https://doi.org/10.1038/s41586-022-05550-y
[6] Cox R, Papoulas O, Shril S … (2026) A protein interactome for the last eukaryotic common ancestor illuminates the biochemical basis of modern genetic diseases. Cell Genomics, 2026; 6
[7] Koumandou, V. L., Wickstead, B., Ginger, M. L., van der Giezen, M., Dacks, J. B., & Field, M. C. (2013). Molecular paleontology and complexity in the last eukaryotic common ancestor. Critical Reviews in Biochemistry and Molecular Biology, 48(4), 373–396. https://doi.org/10.3109/10409238.2013.821444
[8] Richards TA, Eme L, Archibald JM, Leonard G, Coelho SM, de Mendoza A, Dessimoz C, Dolezal P, Fritz-Laylin LK, Gabaldón T, Hampl V, Kops GJPL, Leger MM, Lopez-Garcia P, McInerney JO, Moreira D, Muñoz-Gómez SA, Richter DJ, Ruiz-Trillo I, Santoro AE, Sebé-Pedrós A, Snel B, Stairs CW, Tromer EC, van Hooff JJE, Wickstead B, Williams TA, Roger AJ, Dacks JB, Wideman JG. Reconstructing the last common ancestor of all eukaryotes. PLoS Biol. 2024 Nov 25;22(11):e3002917. doi: 10.1371/journal.pbio.3002917. PMID: 39585925; PMCID: PMC11627563.
[9] Sagan, L. On the origin of mitosing cells. Journal of Theoretical Biology, 14(3), 225–274 (1967).
[10] Tobiasson, V., Luo, J., Wolf, Y.I. et al. Dominant contribution of Asgard archaea to eukaryogenesis. Nature 650, 141–149 (2026). https://doi.org/10.1038/s41586-025-09960-6
[11] Eme, L. et al. Inference and reconstruction of the heimdallarchaeial ancestry of eukaryotes. Nature 618, 992–999 (2023). https://doi.org/10.1038/s41586-023-06186-2