


Symbiose et évolution : à l’origine de la cellule eucaryote
PDF
La cellule des organismes eucaryotes (animaux, plantes, champignons) se distingue de celle des organismes procaryotes (Archées et Bactéries) par la présence de différents organites spécialisés, comme le noyau (contenant l’information génétique de la cellule), la mitochondrie (siège de la respiration cellulaire), ou encore le chloroplaste (siège de la photosynthèse chez les végétaux). L’existence et l’organisation de l’ADN des mitochondries et les chloroplastes, ainsi que leur biochimie et certains traits structuraux, ont conduit à les considérer comme d’anciennes bactéries intégrées dans une cellule hôte par un processus d’endosymbiose. Une des hypothèses possibles serait que les eucaryotes actuels descendraient d’un ancêtre Archée ayant acquis une protéobactérie, qui est devenue la mitochondrie. Une fois cette étape établie, certaines cellules auraient, dans certains cas, intégré des cyanobactéries à l’origine du chloroplaste. Du même coup, elles ont acquis la capacité d’effectuer la photosynthèse, et donc un métabolisme autotrophe. Tout au long du processus, des phénomènes de transferts de gènes entre symbiote, la prise de relai par le noyau du codage de certaines protéines des organites et la relocalisation des produits des gènes dans les organites ont étroitement intégré ces procaryotes au sein de la cellule hôte. Le phénomène d’endosymbiose est donc très largement responsable de la biodiversité des eucaryotes apparus au cours de l’évolution. Ainsi, la photosynthèse s’est développée dans une grande diversité d’organismes : algues rouges et vertes, plantes vertes grâce à des endosymbioses primaires, algues brunes et de très nombreux autres organismes, grâce à des endosymbioses secondaires ou tertiaires.
1. La cellule eucaryote est une chimère
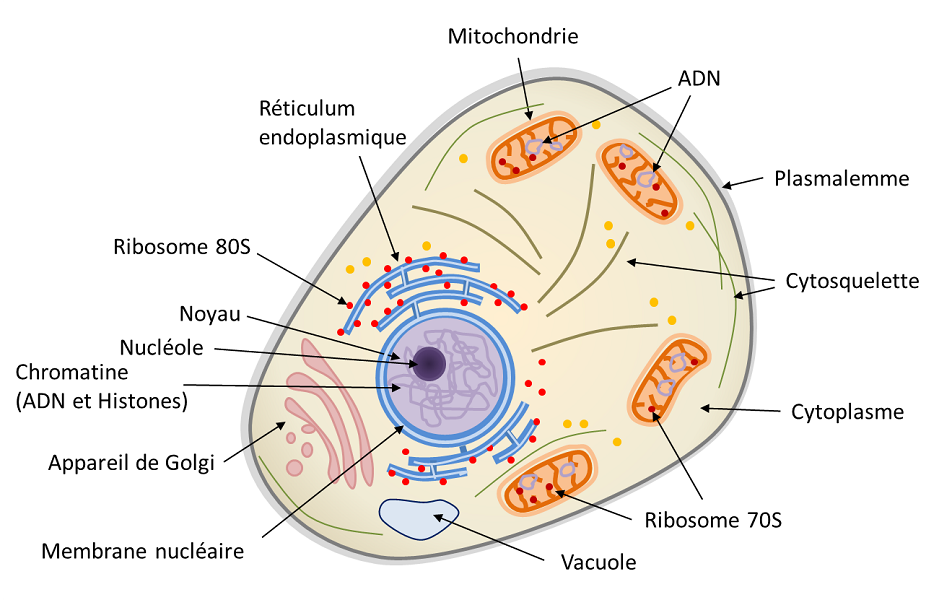
Les eucaryotesOrganismes unicellulaires ou multicellulaires dont les cellules possèdent un noyau et des organites (réticulum endoplasmique, appareil de Golgi, plastes divers, mitochondries, etc.) délimités par des membranes. Les eucaryotes sont, avec les bactéries et les archées, un des trois groupes du vivant. correspondent aux organismes multicellulaires (animaux, plantes, champignons) ainsi qu’à quelques organismes unicellulaires (protozoaires, par exemple). La caractéristique principale de la cellule eucaryote (Figure 1) est l’existence d’un noyau (chez les procaryotes, le génome n’est que très rarement entouré par une membrane) baignant dans un cytoplasme contenant de nombreux organites, comme les mitochondriesOrganites du cytoplasme des cellules eucaryotes (plantes, algues, animaux). Site de la respiration cellulaire, les mitochondries convertissent l’énergie des molécules organiques issues de la digestion (glucose) en énergie directement utilisable par la cellule (l’ATP) au cours du « cycle de Krebs ». Cette réaction nécessite la présence d’oxygène et libère du CO2, elle joue donc un rôle essentiel dans le cycle du carbone. Les mitochondries ont pour origine un organisme procaryote (α-protéobactérie) intégrées dans des protocellules eucaryotes il y a 2 milliards d’années. (siège de la respiration, présente dans toutes les cellules eucaryotes) et les chloroplastesOrganites du cytoplasme des cellules eucaryotes photosynthétiques (plantes, algues). Site de la photosynthèse, les chloroplastes produisent du dioxygène O2 et jouent un rôle essentiel dans le cycle du carbone : ils utilisent l’énergie lumineuse pour fixer le CO2 et synthétiser de la matière organique. Ils sont ainsi responsables de l’autotrophie des végétaux. Les chloroplastes sont le fruit de l’endosymbiose d’un procaryote photosynthétique (de type cyanobactérie) au sein d’une cellule eucaryote, il y a environ 1,5 milliard d’années. (siège de la photosynthèseProcessus bioénergétique qui permet aux plantes, aux algues et à certaines bactéries de synthétiser de la matière organique à partir du CO2 de l’atmosphère en utilisant la lumière du soleil. L’énergie solaire est ainsi utilisée pour oxyder l’eau et réduire le dioxyde de carbone afin de synthétiser des substances organiques (glucides). L’oxydation de l’eau conduit à la formation du dioxygène O2 que l’on retrouve dans l’atmosphère. La photosynthèse est à la base de l’autotrophie, elle est le résultat du fonctionnement intégré du chloroplaste au sein de la cellule., chez les végétaux au sens large, plantes terrestres et algues). Ces organites sont fréquemment déplacés ou réorganisés par le cytosquelette qui anime une mobilité intracellulaire (Figure 1).
Le noyau eucaryote est délimité par une double membrane appelée enveloppe nucléaire (Figure 1). Il contient le génome nucléaire caractéristique de la cellule eucaryote, c’est-à-dire le matériel génétique d’un individu codé dans son ADN (acide désoxyribonucléique). C’est généralement de ce génome dont on parle lorsqu’on mentionne le génome d’un eucaryote. Pourtant, la cellule eucaryote contient aussi des génomes non-nucléaires, au sein des organites :
– le génome mitochondrial, au sein de la matrice des mitochondries (Figure 1) ;
– le génome chloroplastique, au sein du stroma des chloroplastes (cas des plantes ou des algues, par exemple).
L’ADN constitutif de ces trois génomes n’est pas organisé de la même manière. Dans le noyau, le génome est réparti sur plusieurs molécules d’ADN, linéaires et organisées en chromosomes bien différenciés. L’ADN contient toutes les séquences codantes (transcrites en ARN messagers, et traduites en protéines) et non codantes (non transcrites, ou transcrites en ARN, mais non traduites). La configuration tridimensionnelle du génome nucléaire a une importance fonctionnelle : l’enroulement (ou « condensation ») de l’ADN sur lui-même et autour de protéines, les histonesProtéines basiques s’associant à l’ADN pour former la structure de base de la chromatine. Les histones jouent un rôle important dans l’empaquetage et le repliement de l’ADN., permet d’empaqueter une grande quantité d’information génétique dans le minuscule noyau d’une cellule. L’ADN mitochondrial ou chloroplastique n’a pas du tout la même organisation : il est en général circulaire, rarement linéaire (mitochondries de plantes), généralement sans intron, et n’est pas associé à des protéines de type histones.
Les cellules de type procaryoteMicro-organismes (en général unicellulaires) à la structure cellulaire simple, ne comportant pas de noyau, et presque jamais de compartimentation interne (la seule exception étant les thylakoïdes chez les cyanobactéries). Deux des trois groupes constituant le vivant sont des procaryotes : les Archées et les Bactéries. (Bactéries et ArchéesMicroorganismes unicellulaires procaryotes vivant en particulier dans des milieux extrêmes (anaérobies, à forte salinité, très chauds…). Les recherches phylogénétiques de Carl Woese et George E. Fox (1977) ont permis de différencier les archées des autres organismes procaryotes (bactéries). Actuellement, on considère que le vivant est constitué de trois groupes : les archées, les bactéries et les eucaryotes.), quant à elles, ne possèdent pas de noyau et leur ADN est circulaire (ou rarement linéaire) et organisé comme celui des chloroplastes ou des mitochondries. De cette manière la réplication, la transcription et la traduction de l’ADN se fait directement dans le cytoplasme. Il faut cependant souligner que les Archées ne sont que superficiellement similaires aux Bactéries par leur aspect cellulaire : leur métabolisme diffèrent grandement, et les mécanismes et les protéines impliquées dans les processus de réplication, de transcription et de traduction présentent des traits similaires à ceux des eucaryotes. Enfin, les procaryotes n’ont pas toujours de compartimentation interne et les compartiments sont moins complexes (les cyanobactéries sont un exemple d’exception). Surtout, les compartiments, lorsqu’ils existent, ne sont pas mobiles dans la cellule : le cytosquelette, qu’on commence à découvrir, n’y déplace pas les composants cellulaires.
Tableau 1. Comparaison cellules de type eucaryote et procaryote
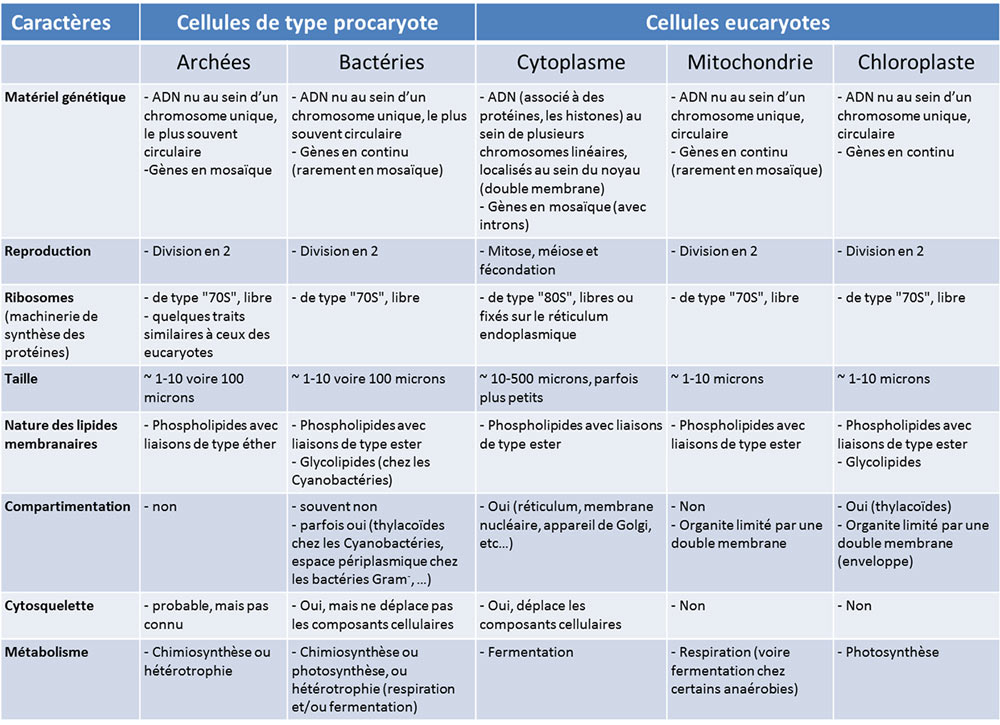
Le Tableau 1 compare les propriétés des cellules procaryotes et eucaryotes (avec leurs mitochondries et éventuellement leurs chloroplastes). Il montre que les mitochondries et les chloroplastes ont de nombreuses caractéristiques communes avec celles des cellules procaryotes. Au-delà de la structure de l’ADN, les organites de la cellule eucaryote se forment à partir d’organites préexistants, se divisant en deux pour se multiplier, à la façon de bactéries. De même, ils disposent de la même machinerie de synthèse des protéines (ribosomesEnorme complexe composé d’ARN et de protéines ribosomiques permettant la traduction des ARNm en protéines. Commun à toutes les cellules (procaryotes et eucaryotes), le ribosome varie en fonction des organismes : ribosome 80S chez les eucaryotes et ribosome de 70S chez les procaryotes et les organites cellulaires (mitochondrie, chloroplaste). de type 70S, libres dans la matrice ou le stroma) alors que dans le cytoplasme de la cellule eucaryote, cette machinerie est constituée de ribosomes de type 80S, parfois fixée sur les membranes du réticulum endoplasmiqueRéseau membranaire du cytoplasme des cellules eucaryotes, essentiel au métabolisme cellulaire (synthèses de lipides et de protéines, stockage du calcium). Associé aux ribosomes, c’est le lieu de synthèse d’une part des protéines secrétées à l’extérieur de la cellule et, d’autre part, des protéines et des lipides constituant les membranes des organites cellulaires (appareil de Golgi, lysosomes, mitochondries, noyau, ribosomes, vésicules…).. Enfin, on retrouve chez des bactéries le métabolisme particulier des mitochondries (la respiration) et des chloroplastes (la photosynthèse). Par contre, la cellule eucaryote se distingue par l’existence d’un réseau protéique actif, le cytosquelette, système auto-organisé et capable de mobilité, qui positionne et déplace les organites dans la cellule. Un tel réseau protéique est statique, voire absent, chez les procaryotes, et peu développé dans les mitochondries et les chloroplastes.
L’ensemble des ces propriétés montre que la cellule eucaryote est une chimère comportant à la fois des constituants caractéristiques de la cellule eucaryote (le noyau) et des organites aux propriétés typiquement procaryotes (chloroplastes, mitochondries).
La distinction entre Procaryotes et Eucaryotes a été proposée en 1925 par le pastorien Edouard Chatton [2] (qui a nommé ces deux types cellulaires), même si elle ne commença d’être reconnu que dans les années 50-60, et la nature chimérique des cellules eucaryotes avait été entrevue dès le tournant du 19e au 20e siècle. Si le botaniste Andreas Schimper (né en France) eu l’idée, en 1883, que les organismes photosynthétiques étaient le résultat de la combinaison d’organismes distincts, ce fut le biologiste Russe Constantin Mereschkowsky, qui le premier apporta des arguments solides pour dire que certaines cellules proviennent d’une union intracellulaire de deux types différents de cellules (endosymbiose). Dans son article de 1905 [3], Mereschkowsky propose trois idées essentielles : (a) les chloroplastes sont des sortes de cyanobactéries qui très tôt dans l’évolution ont établi une symbiose avec un hôte hétérotrophe, (b) l’hôte qui a acquis les plastes était lui-même le produit d’une symbiose antérieure entre une cellule hôte de type amiboïde, plus grosse, hétérotrophe, et un endosymbionte plus petit, de type microcoque, à l’origine du noyau et (c) l’autotrophie des plantes est entièrement héritée des cyanobactéries. Mereschkowsky n’avait pas envisagé l’origine des mitochondries. Elle est à mettre au crédit du microbiologiste français Paul Portier qui écrit dans un texte en 1918 [4] que « tous les êtres vivants, tous les animaux (…), toutes les plantes (…) sont constitués par l’association, l’emboîtement de deux êtres différents. Chaque cellule vivante renferme (…) des formations que les cytologistes désignent sous le nom de « mitochondries ». Ces organites ne seraient pour moi autre chose que des bactéries symbiotiques, ce que je nomme des symbiotes. » Ces observations n’ont pas retenu plus d’attention que cela de la part des scientifiques, et la théorie tomba en disgrâce, en particulier parce qu’on n’a pas réussi à mettre en culture les plastes et les mitochondries, ce qui au 19e siècle était considéré comme la preuve d’une nature bactérienne [5]. Il a fallu l’apport de nouvelles méthodes d’étude de la cellule utilisant la microscopie électronique, la biochimie et la biologie moléculaire pour que la théorie de l’origine endosymbiotique des organites de la cellule eucaryote soit remise au gout du jour vers 1970 par la microbiologiste américaine Lynn Margulis.
2. Comment est apparue la cellule eucaryote ?
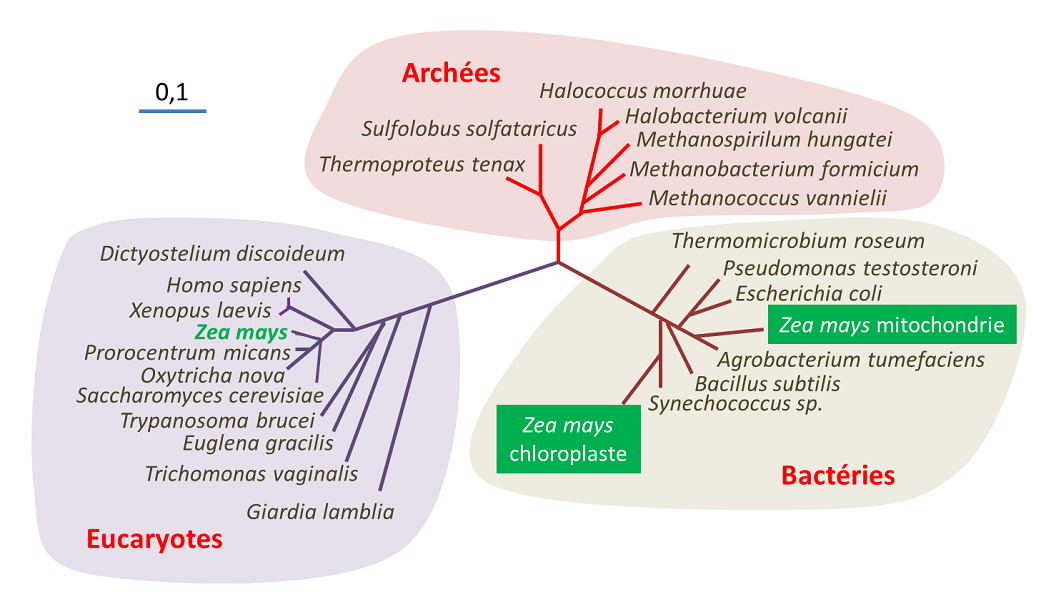
Plusieurs hypothèses ont été émises pour expliquer l’apparition de la cellule eucaryote, il y a environ 1,5 milliards d’années, soit près d’un milliard d’années après l’apparition sur Terre des premiers organismes procaryotes. Cette question peut être abordée de très diverses manières, selon que l’on considère les évidences paléontologiques, les aspects énergétiques, l’origine des caractéristiques de la cellule eucaryote ou les relations des différentes lignées procaryotes et eucaryotes les unes par rapports aux autres [6]. La figure 3 positionne l’origine des eucaryotes et des autres lignées (Archées et Bactéries).
Certains modèles postulent que les eucaryotes ont émergé à partir d’une lignée ancestrale unique via des mutations successives au cours du processus évolutif. D’autres modèles postulent plutôt que les eucaryotes sont apparus à partir d’une association symbiotique de cellules procaryotes dont la fusion aurait réalisé la transition du procaryote à l’eucaryote. Ces diverses hypothèses peuvent en partie être testées par l’expérience, en particulier grâce à l’analyse des génomes des organismes actuels (procaryotes ou eucaryotes) [7,8].
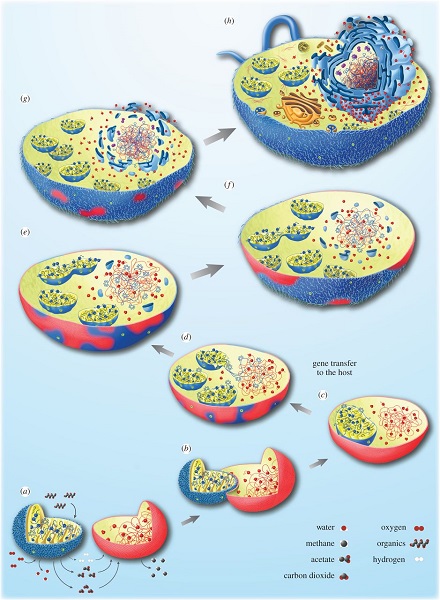
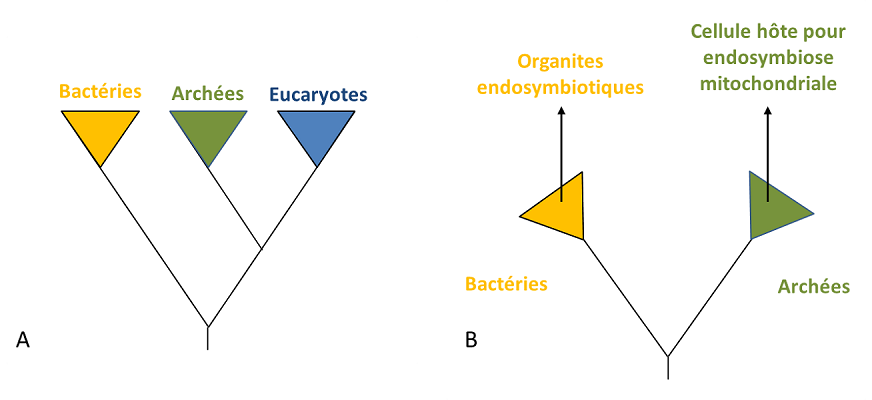
Dans ce contexte, Bactérie et Archée ont tendance à interagir étroitement à la manière de nombreuses associations symbiotiques actuelles. Ceci peut conduire, en principe, à la situation décrite Figure 4, dans laquelle le symbiote bactérien va être retenu par l’hôte archéen et finir par résider à l’intérieur. Dans ce cas, l’hôte ne se nourrit pas du symbiote, aussi l’intégration n’est pas due à un phénomène de phagocytoseProcessus permettant à une cellule d’englober puis de digérer une substance ou organisme étranger (bactérie, par exemple). – même s’il ne fait aucun doute que la phagocytose a augmenté la fréquence d’intégration des endosymbiotes dans la suite de l’évolution des cellules eucaryotes [5].
La question de l’origine de la cellule eucaryote est aussi liée à celle du noyau, structure emblématique de cette cellule. La mise en place d’un nouveau système membranaire, la membrane nucléaire, dans l’hôte après l’acquisition de la mitochondrie pourrait être due à l’agrégation de vésicules membranaires constituées de lipides bactériens. Cette séparation entre noyau et cytoplasme aurait pu répondre à la nécessité de séparer, suite au transfert de gènes entre l’hôte et le symbiote, l’épissage des ARN de la traduction de l’ADN. C’est alors la pression sélective qui aurait conduit à la fixation de la compartimentation entre noyau nouvellement formé et cytoplasme [5] (Figure 4).
Ainsi, tous les eucaryotes actuellement connus descendraient d’un ancêtre Archée ayant acquis au cours du Précambrien une protéobactérie, devenue la mitochondrie. Cette étape est cruciale : l’intégration de la mitochondrie est donc indissociable de l’apparition de la cellule eucaryote telle qu’on la connaît à présent. Les fortes contraintes énergétiques exercées sur l’organisation des cellules procaryotes ont été un facteur d’innovation majeur à l’origine de l’évolution de cette cellule : seules les cellules qui possédaient des mitochondries avaient les ressources énergétiques suffisantes pour atteindre la complexité de la cellule eucaryote, raison pour laquelle il n’y a pas de véritables intermédiaires dans la transition des procaryotes vers les eucaryotes. On considère souvent que ce n’est qu’une fois cette étape établie que certaines de ces cellules auraient acquis les caractéristiques de la cellule eucaryote (noyau, compartimentation) puis, dans certains cas, intégré des cyanobactéries. Du même coup, elles ont acquis la capacité d’effectuer la photosynthèse, à l’origine du chloroplaste, leur conférant ainsi un métabolisme autotrophe.
Récemment, des travaux ont nuancé l’ancienneté de la symbiose mitochondriale [10]. Ils reposent sur l’ancienneté d’acquisition des gènes d’origine bactérienne présents dans le noyau eucaryote (c’est-à-dire, la date de leur divergence avec les gènes homologues qui se trouvent actuellement dans des bactéries libres). Ils ont révélé que de nombreux gènes, dont certains contribuent à la complexité de la cellule eucaryote, avaient été acquis selon toute vraisemblance avant la mitochondrie. Cela n’implique pas que la cellule dans laquelle a pénétré la mitochondrie était aussi complexe qu’actuellement, mais il se peut qu’elle ait par exemple déjà été capable de phagocytose. Ce trait (propre aux eucaryotes car il dépend de la mobilité du cytosquelette) aurait pu aider à la mise en place de la mitochondrie. L’élaboration de la complexité eucaryote reste donc spéculative, mais a pu commencer avant la mitochondrie, même si elle en a ensuite indubitablement profité.
3. L’origine endosymbiotique du chloroplaste
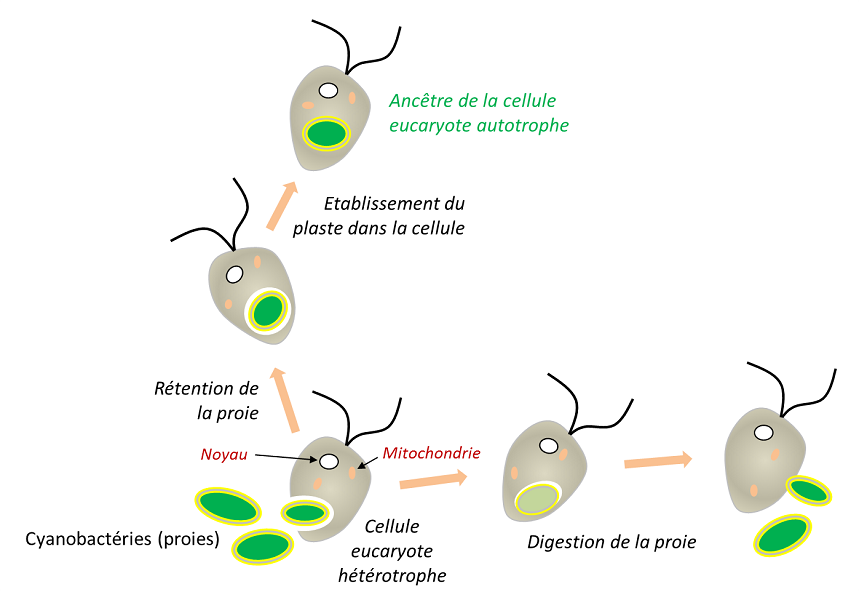
Lors de la phagocytose que l’on observe chez les globules blancs ou de nombreux protozoaires (Figure 5), les cellules ingérées sont souvent directement digérées (c’est le cas des proies), mais il arrive qu’elles soient durablement hébergées dans les cellules (endosymbiotes). Dans le processus d’endosymbiose, l’organite résulte donc de l’internalisation par phagocytose sans digestion d’un procaryote au sein d’un eucaryote (Figure 5). C’est le cas des chloroplastes des végétaux terrestres, mais aussi des algues rouges et vertes qui en sont proches [11,12].
Endosymbioses primaires et secondaires
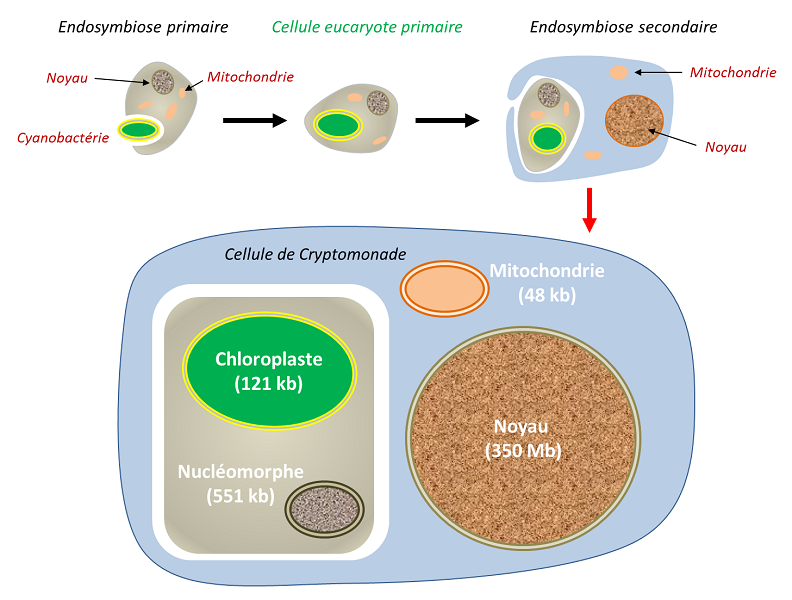
Au cours de l’évolution, plusieurs événements d’endosymbioses se sont répétés et ont conduit à la formation d’organismes particuliers. Lors d’une endosymbiose primaire, la cellule eucaryote intègre un procaryote vivant. Ainsi, les chloroplastes des végétaux de la lignée verte (algues rouges et algues vertes, auxquelles se rattachent les végétaux terrestres) sont issus d’endosymbioses primaires impliquant une cyanobactérie. Chez certains eucaryotes, les mitochondries ont évolué par suite d’une adaptation à des milieux anaérobies, sans jamais disparaître : elles ont engendré des mitochondries particulières (les hydrogénosomesOrganite produisant de l’Hydrogène, dérivé d’une mitochondrie. On en trouve chez certains ciliés anaérobies, les Trichomonas, et les champignons.) réalisant une fermentation productrice de H2 (par exemple chez certains Ciliés) [14], mais aussi des organites de petite taille, seulement impliqués dans des biosynthèses pour la cellule hôte, les mitosomesOrganite présent chez certains organismes unicellulaires eucaryotes, probablement dépourvu d’ADN mais doté de fonctions de biosynthèse. [15].
4. L’intégration du procaryote au sein de la cellule eucaryote
Toutes ces lignées présentent une caractéristique commune : une forte régression génétique des endosymbiotes. Par comparaison avec des protéobactéries libres comme Escherichia coli, les mitochondries ont perdu 99 % de leurs gènes. À l’extrême, les hydrogénosomes et les mitosomes n’ont tout simplement… plus de génome ! Les plastes de la lignée verte révèlent une régression génétique de l’ordre de 95 % par rapport aux cyanobactéries unicellulaires libres : le nombre de gènes est passé de plusieurs milliers chez les cyanobactéries à environ 100 à 200 dans les chloroplastes… voire aucun dans les plastes régressés de la plante parasite Rafflesia.
À l’origine de cette régression figure évidemment la perte des gènes nécessaires à la vie libre, voire à certaines fonctions métaboliques. Par exemple, comme chez toutes les bactéries Gram–Bactéries mises en évidence par une technique de coloration appelée coloration de Gram : elles apparaissent alors roses au microscope. La technique de coloration repose sur les caractéristiques membranaires et de paroi de la bactérie. Néanmoins, il ne s’agit pas d’un facteur de classement phylogénétique : en effet, les groupes ‘Gram +’ et ‘Gram -‘ sont tous deux non-monophylétiques., une couche de peptidoglycaneComposé de la paroi des bactéries à Gram positif et à Gram négatif. Constitué d’une partie glucidique (= polysaccharide) et d’une partie peptidique. Il maintient la forme des cellules et assure une protection mécanique contre la pression osmotique. se trouve entre les deux membranes des cyanobactéries, essentielle au maintien de la structure des bactéries au sein du milieu naturel, de faible osmolaritéNombre de moles de particules « osmotiquement actives » en solution dans 1 litre de solution. Notion reliée à la pression osmotique exercée par les particules en solution, et responsable de l’osmose. Le saccharose est une petite molécule osmotiquement active alors que l’amidon est un énorme polymère de glucose osmotiquement inactif. L’accumulation du saccharose dans un compartiment entraine une augmentation de la pression osmotique dans ce compartiment, ce qui n’est pas le cas de l’amidon.. Une fois intégré au sein de la cellule hôte, le procaryote va se retrouver dans un milieu, le cytoplasme, dont l’osmolarité est très proche de celle de son milieu intérieur. La couche de peptidoglycane devient alors inutile, et les gènes responsables de la mise en place de couche de peptidoglycane sont perdus dans les chloroplastes (sauf chez les glaucophytesEucaryotes unicellulaires du plancton vivant dans les lacs, les mares ou les marécages des régions tempérées. Ils présentent des flagelles (2 de longueur inégale). Ils ont des chloroplastes (appelés cyanelles) de couleur bleu-vert, à cause de la présence de phycocyanine et d’allophycocyanine dans les phycobilisomes. C’est un groupe de diversité réduite.).
Bien que le génome des organites régresse, le répertoire des protéines des organites (le protéome), lorsqu’il est connu, reste semblable à celui du protéome des bactéries libres : des protéines opérant des fonctions nouvelles ont donc compensé les pertes. Leur codage a en fait été pris en charge par le génome nucléaire de l’hôte : des gènes localisés dans le noyau sont traduits dans le cytosol en protéines qui sont adressées à l’organite par le biais d’un peptide de transitSéquence peptidique située à l’extrémité NH2-terminale des protéines nouvellement synthétisées dans le cytoplasme et qui permet de les adresser à l’organite spécifique (mitochondrie, etc…) où elles fonctionnent. On parle aussi de peptide d’adressage.. Ce phénomène de relocalisation dans l’organite du produit des gènes est un phénomène absolument essentiel à l’intégration du procaryote au sein de la cellule hôte. La machinerie d’adressage responsable de ces transferts est une innovation convergente dans les plastes et les mitochondries. Elle constitue d’ailleurs un exemple des fonctions nouvelles liées à la vie intracellulaire. Ces machineries qui permettent l’import les protéines synthétisées dans le cytosol à travers les deux membranes limitantes des mitochondries et des chloroplastes renferment un grand nombre de protéines dont l’origine évolutive est complexe : on y trouve aussi bien des protéines d’origine procaryote que d’origine eucaryote, codées dans l’organite et le noyau. Ensemble, elles permettent la reconnaissance de la protéine en cours d’adressage, son dépliement suivi de l’import (la protéine doit être maintenue dans un état non replié pour traverser les membranes), puis le clivage du peptide d’adressage avant la localisation précise dans son compartiment fonctionnel [19].
Quelle est l’origine des gènes qui codent dans le noyau des fonctions pour les organites ? Il y en a en fait deux (Figure 7) [16]. Parfois, des gènes nucléaires d’origine se sont substitués aux gènes de l’organite : leur produit a acquis la capacité d’être adressé dans l’organite. Cette activation a pu, par le passé, conduire à une situation de redondance chaque fois qu’un gène codait déjà la même fonction dans l’organite. De cette redondance, le gène de l’organite a pu être perdu sans dommage (Figure 7a) [16].
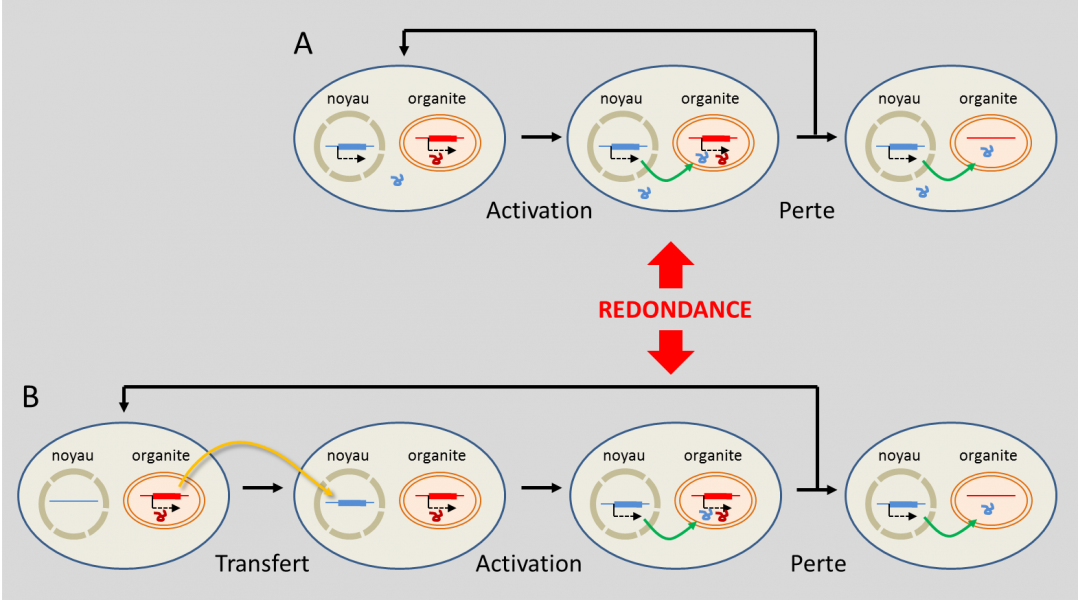
Le passage de fragments d’ADN des organites vers le noyau n’est pas rare : de grands blocs d’ADN des organites sont insérés dans le génome de certaines plantes. Ceux-ci peuvent s’activer : près de 10 % des gènes nucléaires d’Arabidopsis thaliana sont ainsi issus de transferts à partir du plaste, souvent suivis de duplications [20]. On ignore comment l’ADN de l’endosymbiote a pu être intégré dans le génome de l’hôte, mais l’on imagine que cela se produit lors de dégradations d’organites endommagés ou âgés libérant accidentellement au sein du cytoplasme de l’hôte des morceaux d’ADN qui sont ensuite intégrés de manière aléatoire au sein de l’ADN nucléaire de l’hôte.
Les génomes cytoplasmiques des organites sont à la croisée de forces sélectives variées, les unes favorisant leur régression (comme la nécessité de co-expression de certains gènes), les autres favorisant la persistance de tel ou tel gène dans le génome des organites. Cela pourrait être le cas de la sélection pour une petite taille génomique qui accélère la multiplication des organites et permet une meilleure transmission aux cellules-filles : elle sélectionne notamment le transfert de gènes au noyau. Ce dernier cumule ainsi des potentialités génétiques issues de lignées différentes coexistant avec lui dans la cellule [16]. Ainsi, alors que l’endosymbiose réduit les génomes des endosymbiotes, elle nourrit le génome du noyau hôte, contribuant à sa diversification génétique, et poussant à de plus étroites épousailles l’association endosymbiotique. L’endosymbiose mélange donc les lignées évolutives en présence, par emboîtement mais aussi par chimérisation génétique dans le noyau de la cellule hôte.
Enfin, la transmission verticale de l’endosymbiote à travers les générations est essentielle pour que l’endosymbiose perdure. Les plastes doivent se diviser avant la division de la cellule hôte et doivent être distribués pour moitié dans les deux cellules filles. Si leur division est trop rapide, ils pourraient prendre l’avantage sur la cellule hôte ; au contraire, un faible taux de division pourrait conduire à leur disparition. Dans ce contexte, la mise en place d’une coordination de la division cellulaire et de celle du symbiote a été un élément essentiel du succès de l’endosymbiose. Si la plupart des protéines impliquées dans la division des chloroplastes provient de la machinerie de division cellulaire présente chez les cyanobactéries, quelques protéines sont, semble-t-il, d’origine eucaryote, et toutes sont codées dans le noyau : ceci constitue un moyen pour l’hôte d’exercer un contrôle sur la division du chloroplaste.
5. La symbiose moteur de l’évolution ?

Plus qu’une curiosité biologique, la symbiose est certainement l’un des moteurs les plus puissants de l’évolution du monde vivant. Elle crée très rapidement des organismes chimériques qui peuvent engendrer des lignées nouvelles. Elle rapproche des partenaires et favorise des transferts de gènes massifs qui créent des génomes eux aussi chimériques : le génome nucléaire contient ainsi des gènes eucaryotes, mais aussi des gènes d’origine bactérienne, issus des mitochondries, voire des plastes, avec lesquels il voisine. De tels événements expliquent peut-être les sauts évolutifs majeurs dont l’évolution semble ponctuée, qui ont donné naissance aux grandes lignées du vivant et façonné la diversité biologique actuelle.
Ainsi, renouvelant la vision darwinienne d’une évolution par descendance avec modification, où une espèce est susceptible d’en donner deux, les mécanismes d’endosymbiose nous rappellent que parfois deux espèces, auparavant libres et reconnaissables, fusionnent en une seule. L’homme lui-même peut être envisagé comme une communauté symbiotique extrêmement intégrée, formée du cytoplasme eucaryote et des mitochondries, mais aussi des archées et des bactéries qui peuplent son intestin…
Références et notes
Photo de couverture : © Jacques Joyard
[1] Lang T. et al. (2000) Autophagy and the cvt pathway both depend on AUT9. J Bacteriol 182, 2125-2133.
[2] Chatton E. (1938). Titres et travaux scientifiques (1906-1937). Sette, Sottano, Italy. L’histoire des conditions dans lesquelles Chatton a établi le concept de procaryote et eucaryote est décrite par Sapp J. (2005) The Prokaryote-Eukaryote Dichotomy: Meanings and Mythology, Microbiol Mol Biol Rev. 69, 292–305.
[3] Mereschkowsky C. 1905 Uber Natur und Ursprung der Chromatophoren im Pflanzenreiche. Biol.Centralbl. 25, 593–604 ; traduit par Martin W, Kowallik K. (1999) Annotated English translation of Mereschkowsky’s 1905 paper ‘Uber Natur und Ursprung der Chromatophoren im Pflanzenreiche’. Eur. J. Phycol. 34, 287–295.
[4] Portier P. (1918) Les Symbiotes. Masson (ed.), Paris.
[5] Martin W.F., Garg S. & Zimorski V. (2015) Endosymbiotic theories for eukaryote origin. Phil. Trans. R. Soc. B370, 20140330.
[6] Selosse M.A. (2012). Gloire et disgrâce de la théorie endosymbiotique. La Recherche 468: 92-94.
[7] Archibald J.M. (2014) One plus one equals one: symbiosis and the evolution of complex life. Oxford, UK: Oxford University Press.
[8] McFadden G.I. (2014) Origin and Evolution of Plastids and Photosynthesis in Eukaryotes, Cold Spring Harb.Perspect. Biol. 6, a016105
[9] Martin W. & Müller M. 1998 The hydrogen hypothesis for the first eukaryote. Nature 392, 37–41.
[10] Ettema T.J.G. (2016) Mitochondria in the second act. Nature 531, 39–40doi:10.1038/nature16876
[11] Archibald J.M. & Keeling P.J. (2002) Recycled plastids: a ‘green movement’ in eukaryotic evolution. Trends Genetics 18, 577-584.
[12] Keeling P.J. (2004) Diversity and evolutionary history of plastids and their hosts. Am. J. Bot. 91, 1481-1493.
[13] Douce R., Block M.A., Dorne A.J., Joyard J. (1984) The plastid envelope membranes: their structure, composition, and role in chloroplast biogenesis. Subcell. Biochem. 10, 1-84, Springer US (Ed.)
[14] Selosse M.A. & Loiseaux-de Goër S. (1997) La Saga de l’endosymbiose, La Recherche 296, 36
[15] Embley T.M. & Martin W. (2006) Eukaryotic evolution, changes and challenges. Nature 440, 623–630
[16] Lefèvre T., Renaud F., Selosse M.-A. & Thomas F. (2010). Évolution des interactions entre espèces, in F. Thomas, T. Lefèvre & M. Raymond (ed.), Biologie évolutive, p. 530-613. De Boeck, Paris.
[17] Keeling P.J. (2010) The endosymbiotic origin, diversification and fate of plastids. Phil. Trans. R. Soc. B 365, 729-748
[18] Douglas S. et al. (2001) The highly reduced genome of an enslaved algal nucleus. Nature 410, 1091–1096.
[19] Selosse M.A., Albert B. & Godelle B. (2001) Small is successful: selection for reducing organelle’s genome size favours gene transfer to the nucleus. Trends Ecol Evol 16, 135-141.
[20] Jarvis P. (2004) Organellar Proteomics: Chloroplasts in the Spotlight. Current Biology 14, R317-9. http://www.cell.com/current-biology/references/S0960-9822%2804%2900231-3
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Pour citer cet article : SELOSSE Marc-André, JOYARD Jacques (22 novembre 2024), Symbiose et évolution : à l’origine de la cellule eucaryote, Encyclopédie de l’Environnement. Consulté le 16 avril 2026 [en ligne ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/vivant/symbiose-evolution-lorigine-de-cellule-eucaryote/.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.


