亚马孙河地区是仅次于西伯利亚森林的世界第二大森林所在地,但在生物多样性方面却居世界第一。赤道横穿该地区,区域气候“温暖湿润”。虽受季节节律(旱季、雨季)影响,但森林总保持绿色(常绿)。包括世界第一大河——亚马孙河在内的广阔水网为其提供灌溉。从当地到全球,这一独特而复杂的区域都是许多环境、经济和社会问题的核心,涉及的问题是如此之多,以至于科学研究都投入了巨大的精力[1]。尽管如此,我们对它的了解仍然非常有限。这片森林也成为或多或少一些奇幻故事的主题:《黄金国》(Eldorado)、《绿色地狱》(Green Hell)、《翡翠森林》(emerald forest)、《地球之肺》(lung of the planet);它曾庇护着与世隔绝的人们,可怕的美洲印第安人,有时还有食人族,但事实上,他们常常是受害者,而如今被却视为和平爱好者和生态学家……人们的认知并不都是错的,但需要思考:关于这片森林,我们知道什么?我们需要获得哪些知识以及如何获得?如何利用并补充这些知识,以在这种环境下用最佳方式做出决定和采取行动?这样做有什么好处?这里的居民未来如何?
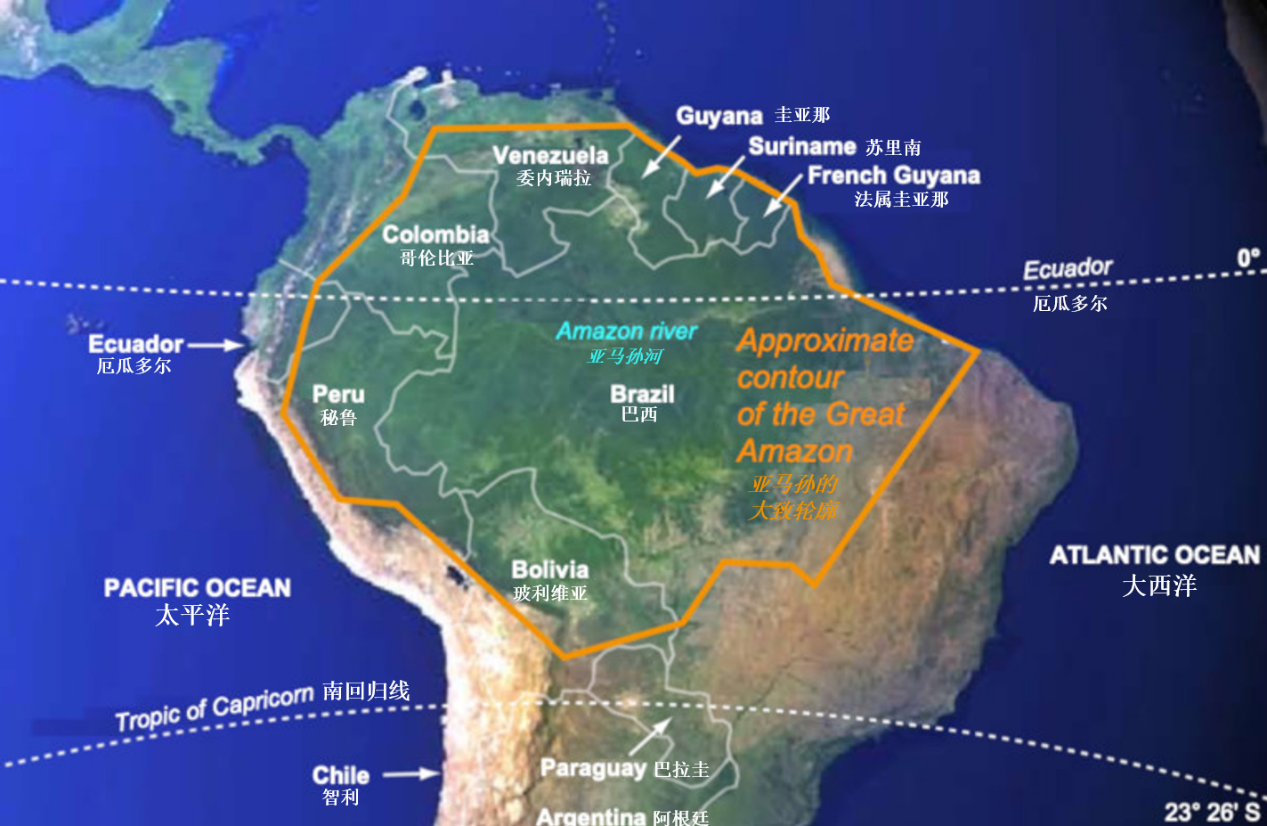
图1. 亚马孙河的部分地理和生物气候边界。森林区域为深绿色。地图参考《环球百科全书》。[来源:Alain Pavé & Gaëlle Fornet, 2010,见参考文献3] (图中,右上赤道上标注“厄瓜多尔(Ecuador)”,有误,应为“赤道(equator)”;“Approximate contour of the Great Amazon”译为“广阔亚马孙的大致轮廓”——校订者注)
包括黄金在内的矿产资源非常重要。疯狂开采破坏了当地环境,然而,绝大多数淘金活动是暗中进行的,其造成的健康和社会后果是灾难性的。另一方面,有些公司试图尽量减小破坏,并在开采后采取修复,如图11所示。巴西的卡拉加斯(Carajas)矿山(产铁和锰)与阿马帕州的塞拉都纳维(Serra do Navio)矿山都做的很不错。值得庆幸的是,曾备受威胁的伦卡(Renca)(约4.1万平方公里,位于帕拉州和阿马帕州交界处)暂时得到了保护[39]。
4. 生态学研究方法的发展
对亚马孙森林的研究已从探险模式转为生态学和人类学研究模式。探险和数据收集的第一阶段是殖民和新殖民主义背景下博物学家、地质学家、地理学家、人类学家和民族学家所做的工作[40]。随后,人们建立了野外台站。就法属圭亚那而言,从1980年代起,帕拉库和努里格(见图7)及其他一些次要台站都得以建立。与此同时,技术革命逐渐将生态学从“刀绳生态学(opinel knife ecology,piece of string)”转变为“技术生态学(technology ecology)”[41](见焦点“法属圭亚那与生态学新技术”)。之后,科学研究的结果得以高效呈现。
无论面对任何情况,仍然可以认为,随机性*和“灵活多变的”相互作用网络提升了这些森林的恢复力*。应该优先研究产生变异性进而产生多样性的过程,不仅在生态学领域如此,而且在更为广泛的生命科学领域也应如此,从而整合对“秩序与混沌生态学(ecology of order and chaos)”的争论[43]。这是可以追溯到2500年前巴门尼德和赫拉克利特争论的现代形式[44]。
[5] U. Pöschl et al, Rainforest Aerosols as Biogenic Nuclei of Clouds and Precipitation in the Amazon. Science, 329, 2010.
[6] Stephen P. Hubbell et al. How many tree species are there in the Amazon and how many of them will go extinct? Proc. Nat. Acad. Sci. USA, 105, supp. 1, 2008.
[7] ter Steege H. et al. Hyperdominance in the Amazonia Tree Flora. Science 342, 2013.
[8] Crowther T. W. et al. Mapping tree density at a global scale, Nature 525, 2015, 201-205.
[9] Lewinsohn T.M., Prado P.I. How many species are there in Brazil? Conserv. Biology, 19, 2005, 619-624.
[12] Fernando D.B. Espírito-Santo et al. Size and frequency of natural forest disturbances and the Amazon forest carbon balance. Nature Communications volume 5, Article number: 3434, 2014
[13] Lie Fan et al. Satellite-observed pantropical carbon dynamics. Nature Plants, 07/29/2019. And “La biomasse aérienne de la végétation de la zone tropicale n’a plus d’impact positif sur le stockage du carbone”
[14] Nascimento L.A. Stockage du carbone dans les sols et dynamique des paysages en Amazonie : l’exemple du Nord-Ouest de l’État de Mato Grosso – Brésil dans le cadre du REDD (Réduction des Émissions par Déforestation et Dégradation).Geography. University Rennes 2, 2015. French. https://www.theses.fr/2015REN20028
[15] Fujisaki K., Perrin A. S., Desjardins T., Bernoux M., Balbino L. C. & Brossard M. (2015). From forest to cropland and pasture systems: a critical review of soil organic carbon stocks changes in Amazonia. Global Change Biology, 21 (7), 2773-2786. ISSN 1354-1013
[16] 对荟萃分析更准确的描述可以查阅:Makowski D., Synthétiser les connaissances en agronomie。Notes from the Académie d’agriculture de France, 2017. https://www.academie-agriculture.fr/publications/notes-academiques/n3af-2017-3-note-de-synthese-synthetiser-les-connaissances-en
[18] Hoorn C. & Wesselingh F (Eds). Amazonia Lanscape and Evolution. A look into the past. Whiley-Blackwell, Oxford, UK, 2010.
[19] Randford T., Could humans cause another Paleocene-Eocene Thermal Maximum? Climate Homenews, 2013.
[20] Schwarz D. Expansions et recul des forêts équatoriales. Pour la Science, 271, 2000. 在这篇法语文章中,我们可以找到以下两篇参考文献中介绍的大部分结果。这些结果主要来自由联合国教科文组织(UNESCO)资助、IRD/CNRS(环境项目)发起的ECOFIT“热带森林生态系统”项目。
[21] Poncy O., Sabatier D., Prévost M.F. & Hardy I. The lowland high rainforest structure and tree species diversity, in Bongers F., Charles-Dominique P., Forget P.M. & Théry M. (Eds) ” Nouragues. Dynamics and Plant-Animal Interactions in a Neotropical Rainforest”, Kluwer Acad. 2001, 32-46.
[22] Servant M & Servant-Vildary S. (Eds, 2000). Dynamiques à long terme des écosystèmes forestiers intertropicaux CNRS, UNESCO, MAE, IRD, Paris, 427p.
[23] Pavé A. Necessity of chance: biological roulettes and biodiversity, C.R. Biologies, 330, 2007, pp. 189-198
[25] Gourlé-Fleury S., Guehl J.M. & Laroussinie O. Ecology and Management of a Neotropical Forest. Lessons drawn from Paracou, a long-term experimental research site in French Guiana. Elsevier, 311p, 2004.
[26] Saint-Jean D. & Pellet E. Explorateurs d’Amazonie. Aventuriers de la Science en Guyane. Ibis Rouge Éditions, 2008 (Preface Alain Pavé).
[29] Norten N., Angarita H. A., Bongers F., Martinez-Ramos M., Granzow-de la Cerda I., van Breugel M., Lebrija-Tejos E., Meave J.A., Vandermeer J., Bruce Williamson G., Finegan B., Mesquita R. & Chazdon R.L. Successional dynamics in neotropical forests are as uncertain as they are predictable. Proc. Nat. Acad. Sci. USA, 112, 8013-8018, 2015.
[30] 安娜·罗斯福(Anna Roosevelt)是研究亚马孙的最著名的考古学家和生态学家之一。从她的曾祖父西奥多·罗斯福(Theodore Roosevelt),这个家族就对这个地区感兴趣。西奥多曾任美国总统(1901—1909),并在两届任期后多次在亚马孙逗留。安娜有许多作品,如:Roosevelt A. Twelve Thousand Years of Human-Environment Interaction in the Amazon Floodplain. Advances in Economic Botany, Vol. 13, New York Botanical Garden. pp. 371-392, 1999. 她是法国国家科学研究中心亚马孙项目科学委员会成员。
[31] Reich D. et al. Reconstructing Native American population history. Nature, 488, 370-375, 2012. 该工作由法国国家科学研究中心亚马孙项目资助。
[32] Rostain S., Amazonie : les 12 travaux des civilisations précolombiennes. Belin, Paris, 2017; & Rostain S., Amazonie un jardin sauvage ou une force domestiquée. Essai d’écologie historique.. Actes Sud, wandering, 2011.
[33] Molino J.F., Mestre M. & Odonne G. La biodiversité de l’Amazonie, un héritage des Précolombiens ? Research, 527, 67-71, 2017.
[34] Jérémie S., Dambrine E.. Impact des occupations amérindiennes anciennes sur les propriétés des sols et la diversité des forêts guyanaises. In, Alain Pavé and Gaëlle Fornet, Op. Cit.
[35] McKey D., Rostain S., Iriarte J., Glaser B., Birk J.J., Holst I. &Renard D., “Pre-Columbian Agricultural Landscapes, Ecosystem Engineers, and Self-organized Patchiness in Amazonia”, Proc. Nat. Acad. Sci. USA, 107, 2010, p. 7823-7828. CNRS press release, March 13, 2010.
[38] http://www.epoc.u-bordeaux.fr/index.php?lang=frage=eq_ea_flash01 and the summary document: https://hist-geographie.dis.ac-guyane.fr/…/2d1_les_defis_de_la_sante_en_guyane.doc
[39] 伦卡(Reserva Nacional do Cobre e Associados)于1980年代早期出于未来采矿的目的成立。在未进行开发的情况下,它实际上成了自然保护区。2017年,时任巴西总统米歇尔·特梅尔(Michel Temer)对这一状态构成威胁。面对国内外抗议,其自然保护区的状态得以保持。
[40] Raby M. The Colonial Origin of Tropical Field Station. Am. Scientist, 105, 216-223, 2017.
[41] Legay J.M. & Barbault R. (Dir.). La révolution technologique en écologie. Masson, 1995.
[42] Pavé A., Comprendre la biodiversité, vrais problèmes et idées fausses. Threshold Edition, 2019.
[43] Worster D. The Wealth of Nature, Environmental History and the Ecological Imagination. Oxford University Press, 1993.
The Amazon region is home to the second largest forest on the planet after the Siberian forest, but the first in terms of biodiversity. The equator crosses it; the regional climate is described as “warm and humid”. It is subject to a seasonal rhythm (dry season, wet season), such that it always remains green (evergreen). A broad hydrographic network irrigates it, including the Amazon, the largest river in the world. This unique and complex object is at the heart of many environmental, economic and social issues, from local to global, so much that scientific research devotes significant efforts to it.[1] Nevertheless, our ignorance about it is still great. This forest is also the subject of more or less fantasized stories: Eldorado, Green Hell, emerald forest, lung of the planet; it would have sheltered isolated peoples, fearsome Amerindians, sometimes cannibals, in fact often victims and nowadays perceived as peaceful and ecologists… Not everything is wrong, but to be put into perspective: what do we know about it? What knowledge should be acquired and how to do it? How can they be used and complemented to decide and act in the best way on this environment? To what benefit? What future for the people who live there?
The Amazonian forest ecosystem is attributed a global role in regulating greenhousegases (GHGs) and as a reservoir of biological diversity. This ecosystem is also an economic, human and social object through the resources it contains, the populations that inhabit it and the attention it arouses. Spread over nine countries, its future depends on various policies, including those of non-Amazonian institutions. Bioclimatic changes are likely to change its functioning. Human actions can alter its structure: from the simple cutting of plots to the deforestation of entire regions, or, more subtly, local changes such as the enrichment of “useful” species. The representations we make of them, the ideas that emerge from them, sometimes far from the realities on the ground, play a decisive role, particularly in political and technical decisions. Finally, spontaneous biophysical and ecological processes give it a highly random structure. This is the case, for example, of the spread of plant seeds, especially those of trees, by fluids, air and water, whose flows are often turbulent, or by animals (birds, mammals) whose movements are mostly erratic.
Figure 1. Some geographical and bioclimatic landmarks of the Amazon. The forest area is in dark green. Map based on the Encyclopædia Universalis atlas. [Source: Alain Pavé & Gaëlle Fornet, 2010, see ref. 3]The word “forest” refers to an ecosystem, densely populated with trees and occupying a defined terrestrial space. There are different types depending on the bioclimatic conditions in which the forests are located. Thus, we can distinguish boreal forests, temperate forests, mediterranean forests, intertropical forests. Subcategories have also been defined, such as temperate or intertropical dry and humid forests. The Amazonian forest is therefore of the “intertropical humid” type.
Beyond these generalities, the data acquired on this ecosystem show significant variations in space and time. These variations are often due to the techniques and methods used by researchers. To these metrological variations, it is necessary to add the variability generated by bioclimatic and ecological processes influencing ecosystem dynamics. Human actions also matter a lot, quite soft and subtle in the past they can be brutal: massive destruction of trees followed by arsons.
The forest is drained by a vast hydrological system acting on forest dynamics and vice versa. The Amazon flow at the estuary is estimated at between 200,000 and 250,000 m3/s, on average over the year; the amount of sediment arriving in the ocean is about one billion tonnes per year, its basin covers an area of 6,790,000 km2. Significant areas are occasionally flooded and transiently constitute large wetlands (Figure 3).
The word “entangled“, used by Charles Darwin (read Focus Darwin) in the last paragraph of “The Origin of Species“, seems appropriate to reflect the complexity of an Amazonian ecosystem, as suggested by the photography at the top of this article.
The global characteristics of forest stands are increasingly well known: 16,000 highly mixed tree species, of which 227 are dominant, a total of 390 billion trees [6],[7]. By way of comparison, the number of trees worldwide is estimated at 3,040 billion (nearly 8 times higher than a previous estimate of 400.25 billion), including 1,390 billion in tropical and subtropical forests, 740 billion in boreal forests and 610 billion in temperate forests. 15 billion would be cut each year and it is estimated that 46% have disappeared since the beginning of human civilizations [8]. It is clear that the differences between estimates over time are not negligible, most talks on this subject do not take these uncertainties into account. In addition, tree cover is highly dependent on climatic conditions, changes over time and is minimal during ice ages.
The Amazonian ecosystem contains 10 to 13% of the continental biodiversity, for 5% of the land surface [9]. There is no precise census of the species present in the Amazon. The one concerning trees is particular [10]. All categories combined, we can take as benchmarks some estimates collected in the literature: of the order of 2 million living species, including, for example, ~45,000 plant species, ~1.3 million animal species, including ~1 million insects, of the order of 500 for mammals, 1,300 for birds and 3000 for fishes in rivers. We are extremely far from having identified everything and even simply having reliable estimates for all groups [11]. Today, research focuses more on the study of processes than on censuses and, nowadays, the latter are only done according to the needs of functional studies.
Nevertheless, the El Niño and La Niña events in the Pacific are having a rapid impact in the Amazon, to the point that over the period 2010-2017 the balance is zero and for what has been assessed in the past, very variable [13]. During El Niño, changes in the Antarctic circumpolar current lead to an increase in dry winds from Antarctica. These mechanisms have made it possible to explain past events, for example, the dry periods of the Holocene (see below). Such sequences are episodic.
On a daytime scale, GHG exchanges vary, the nocturnal respiration of plants releases CO2, but diurnal photosynthesis absorbs it. At the seasonal level, CO2 sequestration is more efficient in the rainy season than in the dry season.
While the carbon stock above ground can be roughly estimated (see above), as far as soils are concerned, difficulties are encountered due to the size of the Amazonian system, soil diversity, vegetation cover, agricultural and other uses, as well as technical and methodological aspects of measuring and exploiting results. A major effort has been made in recent years with the implementation of REDD (Reducing Emissions from Deforestation and Forest Degradation in Developing Countries), resulting in changes in agricultural operations and land use more generally.
It would be bold to propose estimates for the Amazon and its forest. To give an idea of the variability of the results, we can quote the work of Lucelma Aparecida Nascimento [14], which gives estimates ranging from 0 kg/m2 to 8.89 kg/m2 of carbon for forest soils (northwest of Mato Grosso). This work contains many data and analytical results for various soils and vegetation covers. The effects of land-use change are being assessed, and initial results show that pasture soils have a higher organic carbon stock than forests. On the other hand, the opposite is true for cultivated soils[15]. It should be noted that this type of study increasingly uses meta-analysis [16] to complement specific field work.
In any case, the Amazonian forest plays an important role in the major biogeochemical balances. During the day, like all forests, it absorbs carbon dioxide and releases oxygen (owing to photosynthesis). At night, it “breathes” using oxygen and releases CO2 through respiration. It constantly emits other gaseous compounds: NOx, VOCs, etc. It stores carbon mainly in its trees and contributes to maintaining a humid climate at the regional level. However, it is by no means the “lung of the planet” nor the main producer of oxygen, other forests and grasslands contribute to it and especially the oceans. As we will see later, it produces usable renewable resources including wood (a long-term means of carbon storage!). The first to be concerned with the maintenance, or even development, of this production are the inhabitants of this forest, who should be discussed first, before discussing the “global role” (see section 3).
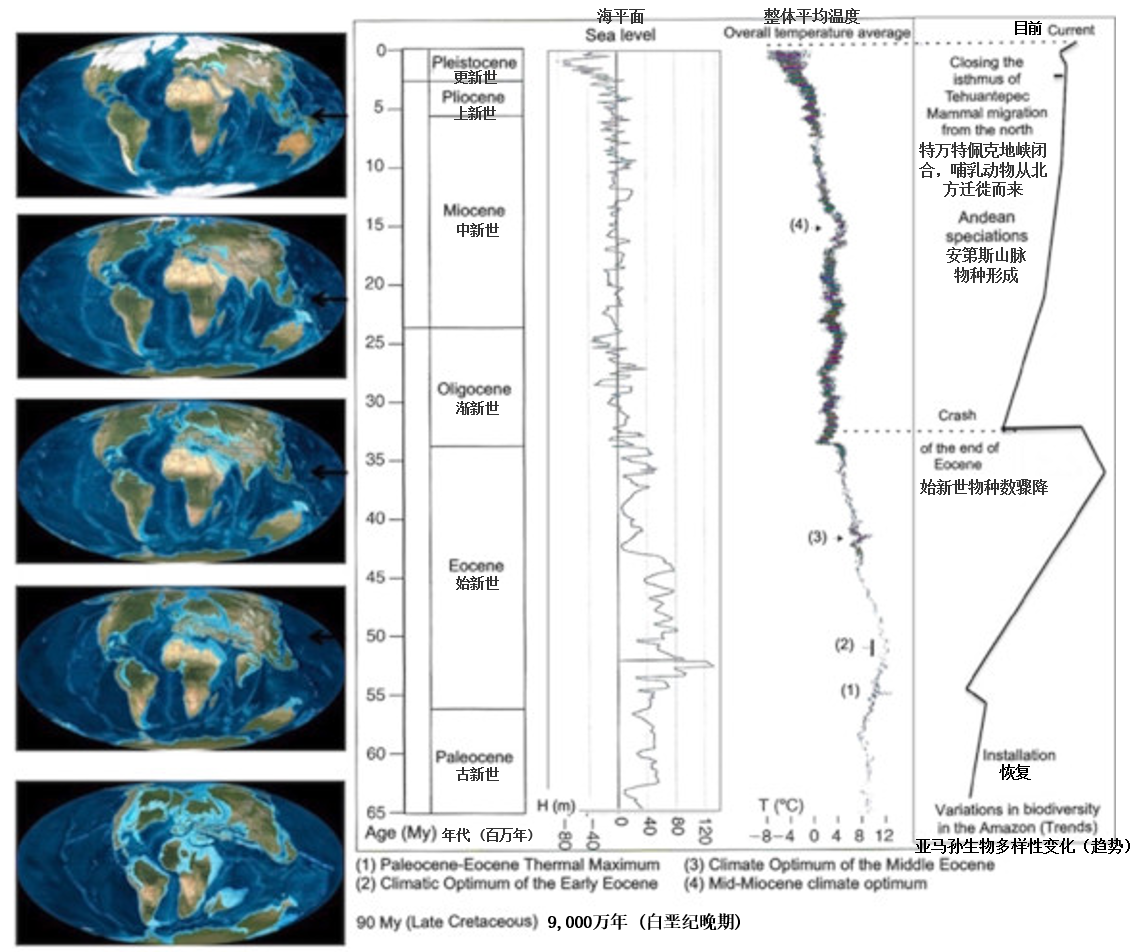
1.4. History
Figure 5. Historical landmarks: variations in biodiversity of the Amazonian system and correlative events, including geological events. [Source: Alain Pavé & Gaëlle Fornet, 2010, see ref. 3]The beginning of the sttlement of this forest is estimated at 55 million years, i.e. in the middle of the Eocene (Figure 5). Its biodiversity was at its highest at the end of this period (around 37 million years BP) [17]. The average temperature on the globe was then 10 to 12°C higher than that of our time [18], but unevenly distributed across the globe and particularly in the Amazon where it was about 5°C higher [19]. The poles had little or no ice, the mountains few or no glaciers. The closure of the Isthmus of Panama 3 million years ago, which allowed the migration of large predators from north to south, explains the decline in diversity in the vicinity of the Pleistocene-Pliocene transition.
During the last glaciation, the Amazonian forest seems to have resisted drought well. Over the past 10,000 years (Holocene), there have been significant variations in climate and forest cover, ranging from 10,000 to 8,000 BP, 6,000 to 4,000 BP and more recently between 1,500 and 1,100 BP and 800 to 500 BP [20],[21],[22].
2. Structure and functioning of the Amazonian forest
This forest is different from other already well-known cases, such as those of French forests:
Natural regenerationversus regeneration mainly carried out by humans;
Bioclimatic zone (inter-tropicalversus temperate);
High biodiversity (one hectare of Guyanese forest contains as many indigenous tree species, on average 200, as in the entire metropolitan territory: 137 for 55 million hectares);
Very different scales (billions versus, at most, thousands of hectares), etc.
What is presented below provides more information while knowing that the major fundamental ecological mechanisms are present.
Figure 6. This map of the Amazonian forest of French Guiana was compiled from remote sensing data. The shades of green correspond to different forest stands. The location of research stations is specified as well as the location of laboratories in Cayenne and Kourou (see ref. [27]). [Source: Alain Pavé & Gaëlle Fornet, 2010, see ref. 3]The distribution of trees is highly random, heterogeneous and mixed: neighbouring individuals are generally of different species (up to 200 species per hectare are represented by at least one individual). It is easy to show that such a structure promotes its resilience and the maintenance of biodiversity[23]. This is observed in the forest plots studied, for example in the forest land stations in French Guiana [24]: Paracou [25] and Nouragues. On a larger scale, heterogeneity is visible from aerial and satellite sensor images, in accordance with an ocean gradient in the continental interior, particularly rainfall, which is reflected in forest types (Figure 6) [26],[27].
Below these broad outlines, there are multiple ecological relationships between living beings, for example: competition between trees (access to light and water resources), predation by animals, cooperation between trees and animals for the dissemination of seeds, a process known as zoochore dominant in the Amazon, or between trees such as the sharing of mycorrhizae*, VOC emissions can signal the arrival of predators or pests (xylophages or tree leaf eaters) [28]. All this reminds us that the forest is not just a collection of trees, but an ecosystem populated by other plants, animals and micro-organisms. These various entities interact.
Spontaneous regeneration is quite well known: after a disturbance corresponding to the destruction of trees, trees of heliophilic species* settle. Then, under the cover thus formed, a rather ombrophilic vegetation gradually replaces the heliophiles. The piece of forest is reconstituted, not identically, but with biodiversity close to that of the neighbouring and previous stand [29].
The Amazonian forest has long been considered primary* or pristine, where humans would have played a negligible, if not non-existent, role in its structure and functioning. In fact, many observations have attracted the attention of researchers, even in the absence of archaeological artifacts, such as local enrichment with “useful” species or sustainable soil transformations. Humans have participated in the dynamics of this forest. It is necessary to revisit the very notions of primary or virgin forests following this observation.
In fact, to start real fires, large areas must be deforested, then allowed to dry and burn. Most often, the fire stops at the edge of the forest, otherwise the progression stops quickly as the humidity is high. However, there is concern that in the event of massive deforestation, the self-sustaining climate regime will stop and cause a chain reaction of drying out and thus fire sensitivity.
The peoples of the forest are Amerindians, but also of “black-brown” origin, i.e. slaves who have escaped from the large plantations. Those who escaped acculturation have retained African knowledge. This is the case for inland navigation and the construction of large canoes necessary for this navigation. Along the major rivers, there are also mixed populations of “caboclos“, mainly from Portuguese and Amerindian marriages. On their side, the majority of inhabitants of European origin are concentrated in urban areas. In total, a human population of great diversity.
The Amazonian living resources are numerous. Some have been appropriated by Europeans and widely valued elsewhere, for example, cocoa, pineapple, cassava, many varieties of beans [36], palm heart, etc. There are many high quality, under-exploited wood resources that could be exploited using modern “sustainable” extraction techniques for drying and machining. Hevea, a latex producer, has been very successful thanks to plantations in many intertropical regions. Balata rubber has remarkable mechanical properties and is used in the manufacture of “top-of-the-range” golf balls. Among the medicines, we can mention quinine, from the quinquinas of the rubiaceae family, found on the Amazonian slope of the Andes. However, we note the limited number of pharmaceutical products from the forest and traditional knowledge for which there are about 1,300 plants for medicinal use but which have not given rise to any real development. Biopiracy for pharmacological purposes is more a myth than a reality. It is probably not the “green gold of the Amazon” [37].
The state of the environment is an important factor to consider in improving the health of Amazonian populations, in addition to pathogens and their vectors, pollution can alter it, such as mercury pollution from gold panning [38]. It should be noted, however, that physico-chemical pollution can be stopped by cutting off the source. It remains at the worst constant, if not gradually decreases. This is not the case for pathogens that reproduce and spread. They also evolve rapidly and can become resistant to control measures, including antibiotics. Biodiversity is not good in essence and we are far from having gone around: plants are frighteningly toxic, animals are dangerous, etc.
Mineral resources, including gold, are important. Their wild exploitation disrupts the local environment, yet most of the gold panning is clandestine, and the health and social consequences are disastrous. On the other hand, some companies try to minimize damage and carry out restoration after exploitation, as shown in Figure 11. In Brazil, the case of the Carajas mines (iron and manganese) is spectacular, as well as the Serra do Navio mine in Amapá. Fortunately, Renca (about 41,000 km2 on the borders of the Para and Amapa states), once threatened, remains protected for the time being [39].
4. Evolving the ecological approach
The study of the Amazonian forest has shifted from an exploratory mode to an ecological and anthropological one. The first phase of exploration and data collection was the work of naturalists, geologists, geographers, anthropologists and ethnologists in a colonial and neo-colonial context [40]. Then field stations were set up; for French Guiana, from the 1980s onwards, these were Paracou and Nouragues (see Figure 7) and others of lesser importance. At the same time, a technological revolution has gradually transformed ecology from “opinel knife ecology, piece of string” to “technological ecology” [41] (see Focus French Guiana and new technologies for ecology). Scientific research can then be expressed effectively.
The stakes are very high, we must manage these forests with various objectives, including maintaining a high level of biodiversity while preserving the interests of the inhabitants. This stewardship must be based on solid basic knowledge and a multivariate approach and not on a one-dimensional projection, for example, on that of the strict economic factor [42]. An ideal context for the design of a true ecological systems engineering.
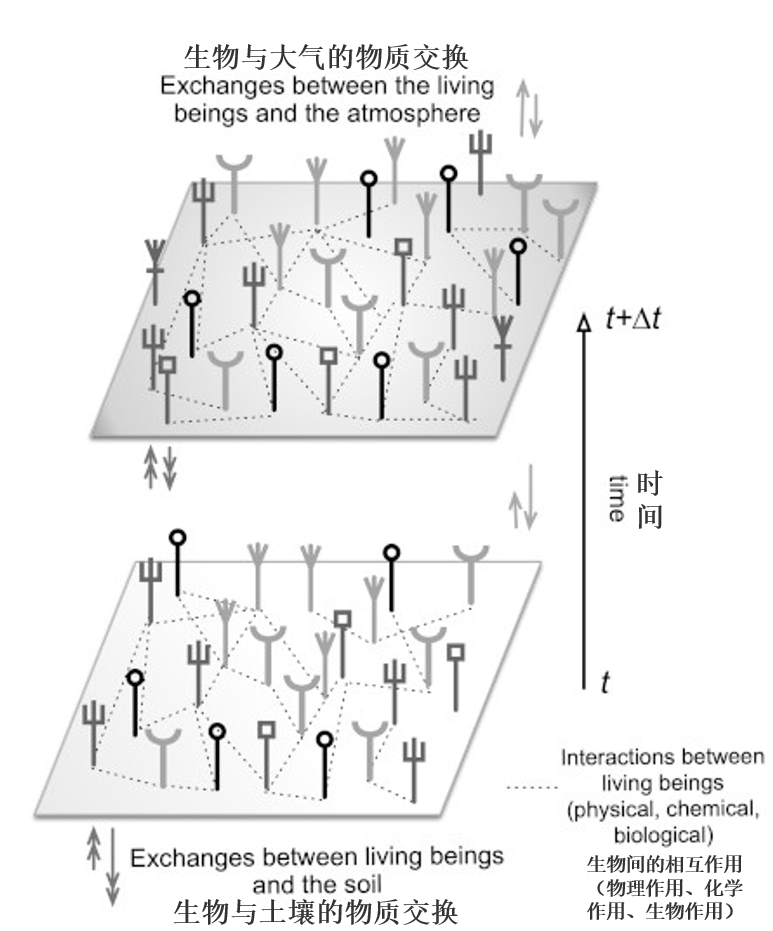
4.1. An Amazonian forest in an unstable state
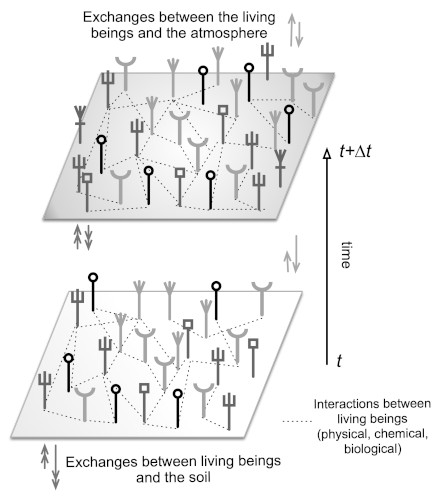
Figure 12. Schematic diagram of a natural forest ecosystem, the trees are represented by vertical lines ending in a different top. These various individuals may interact with each other. In addition, they interact with the physical environment: atmosphere, hydrosphere and soil. These individuals and the various interactions can change over time. They are most often non-linear. [Source: Alain Pavé & Gaëlle Fornet, 2010, see ref. 3]Below an apparent homogeneity of the forest, different stands can be distinguished, even from a distance, with sensors on board satellites. Moreover, this forest is very heterogeneous and mixed; on a small scale, the neighbouring trees are of different species and ages and therefore of different sizes. Ecological communities are also distinguished by edaphic* (drained or undrained soils) and bioclimatic conditions, such as, for example, the ocean-continental gradient in Guyana, visible in Figure 7. The age of the Amazonian forest and its history show that it has not always been the same, it varies permanently, but slowly on a human time scale. Here, the concept of climax* is to be relativized if not abandoned. Punctual observation gives the illusion of balance, which can lead to a fixistic conception of nature. Human action can disrupt this condition as well as natural accidents intermittently. Similarly, interactions between the entities that make up this ecosystem contribute to its evolution.
These interactions are of different natures, between trees themselves or involving other living beings, such as animals or micro-organisms, or simply physical or chemical factors (Figure 12).
The diversity, the number of objects involved and the non-linearity of their interactions lead to the qualification of such a system as complex. Are there “emerging” properties that cannot be reduced to statistical balances? The question is open.
4.2. Chance at the handles
In any case, it can still be argued that stochasticity* and “flexible and changing” interaction networks promote the resilience* of these forests. The study of processes generating variability and therefore diversity should become a priority, not only in ecology, but more generally in life sciences, thus integrating the debate on the “ecology of order and chaos” [43], itself a modern form of the controversy between Parmenides and Heraclitus dating back… 2500 years [44].
A major theoretical effort must be made to move forward, for example on biodiversity, at the risk that the much-publicized erosion may be primarily that of the concept itself. This is the concern of several authors [45], including the one in this article.
Finally, biology has acquired most of its knowledge on a limited number of biological models, roughly organisms of about fifty species including the famous drosophila and Escherichia coli. Some ecosystems could play this role for ecology and all or part of the Amazonian forest could be one of them.
What do we know about it? Despite a major effort over a long period of time, data for the Amazon are still fragmentary and imprecise. The forest is not just a collection of trees, it is an ecosystem with other plants, animals, microorganisms, irrigated by a vast hydrological system, with multiple interactions. Beware of simplistic messages and descriptions that are more poetic than scientific.
What knowledge should be acquired and how to do it? In addition to factual data, develop a synthetic, systemic representation of the region to guide the acquisition of environmental data, which also requires continuing the technological effort. Do not hesitate to conduct theoretical reflections.
How can they be used to decide and act in the best way on this environment? Model this set to test evolution scenarios, promote adaptive household and stewardship strategies, and define a real engineering of ecological systems.
To what benefit? First of all, to the inhabitants of this region, but also to the nations that shelter it, without forgetting the global role of the Amazonian ecosystem (climate and biodiversity), especially the forest, which concerns humanity more broadly.
What future for the people who live there? (Figure 13) This future must be “reasoned, chosen and shared”, constantly reassessed and adapted, by implementing dynamic management accompanied by modelling and simulation [46].
Notes and references
Cover image. [Source: CNRS, Pavé A. & Fornet G. ref. 3]
[1] The French contribution is mainly made in French Guiana, in collaboration with teams from other countries, notably Brazil.
[2] Over time, sources of error are identified and corrected. This explains the variability of the estimates, 5.5 million km2 is a minimal estimate. General reminder: world area = 510 million km2 (510.067,420), oceans = 362 million km2 (361.8) and land area = 148 million km2 (148.2), forest area (2010) = 4.033 billion ha or 40.033 million km2 (27% of land area).
[3] Pavé A. & Fornet G., Amazonie, une aventure scientifique et humaine du CNRS, Ed. Galaade, Paris. 2010. (in french)
[4] Radiation whose wavelengths range from 400 to 700 nm, causing photosynthesis in plants. The light energy received by the illuminated surface is expressed in µmol.m-2.s-1.
[5] U. Pöschl et al, Rainforest Aerosols as Biogenic Nuclei of Clouds and Precipitation in the Amazon. Science, 329, 2010, 1513-1516; Huisl W. Pöschl U., The Amazon rainforest – a cloud factory. Max Planck Gesellschaft. 2010.
[6] Stephen P. Hubbell et al. How many tree species are there in the Amazon and how many of them will go go extinct? Proc. Nat. Acad. Sci. USA, 105, supp. 1, 2008.
[7] ter Steege H. et al. Hyperdominance in the Amazonia Tree Flora. Science 342, 2013.
[8] Crowther T. W. et al. Mapping tree density at a global scale, Nature 525, 2015, 201-205.
[9] Lewinsohn T.M., Prado P.I. How many species are there in Brazil? Conserv. Biology, 19, 2005, 619-624.
[10] Indeed, the trees remain in place. They are large in size. They have a long life span. Their observation is easier than that of mobile and smaller organisms, such as animals. However, trees move, but the movement is intergenerational: seed dispersal, followed by success or failure of germination and growth according to local bio-ecological conditions. This is how tree populations can migrate during bioclimatic variations.
[11] In fact, new species are regularly discovered in the Amazon. In a WWF report published on 30 August 2017, 381 new species were identified, excluding insects.
[12] Fernando D.B. Espírito-Santo et al. Size and frequency of natural forest disturbances and the Amazon forest carbon balance. Nature Communications volume 5, Article number: 3434, 2014
[14] Nascimento L.A. Stockage du carbone dans les sols et dynamique des paysages en Amazonie : l’exemple du Nord-Ouest de l’État de Mato Grosso – Brésil dans le cadre du REDD (Réduction des Émissions par Déforestation et Dégradation).Geography. University Rennes 2, 2015. French. https://www.theses.fr/2015REN20028
[15] Fujisaki K., Perrin A. S., Desjardins T., Bernoux M., Balbino L. C. & Brossard M. (2015). From forest to cropland and pasture systems: a critical review of soil organic carbon stocks changes in Amazonia. Global Change Biology, 21 (7), 2773-2786. ISSN 1354-1013
[16] For more precision on the meta-analysis, we will be able to consult: Makowski D., Synthétiser les connaissances en agronomie. Notes from the Académie d’agriculture de France, 2017. https://www.academie-agriculture.fr/publications/notes-academiques/n3af-2017-3-note-de-synthese-synthetiser-les-connaissances-en
[17] BP (Before Present), BC (Before Christ), AC (After Christ).
[18] Hoorn C. & Wesselingh F (Eds). Amazonia Lanscape and Evolution. A look into the past. Whiley-Blackwell, Oxford, UK, 2010.
[20] Schwarz D. Expansions et recul des forêts équatoriales. Pour la Science, 271, 2000.
In this article (in French), we find most of the results presented in the following two references and obtained mainly by the ECOFIT programme “Inter-tropical Forest Ecosystems”, an IRD/CNRS (Environment Programme) initiative, supported by UNESCO.
[21] Poncy O., Sabatier D., Prévost M.F. & Hardy I. The lowland high rainforest structure and tree species diversity, in Bongers F., Charles-Dominique P., Forget P.M. & Théry M. (Eds) ” Nouragues. Dynamics and Plant-Animal Interactions in a Neotropical Rainforest”, Kluwer Acad. 2001, 32-46.
[22] Servant M & Servant-Vildary S. (Eds, 2000). Dynamiques à long terme des écosystèmes forestiers intertropicaux CNRS, UNESCO, MAE, IRD, Paris, 427p. (in French)
[23] Pavé A. Necessity of chance: biological roulettes and biodiversity, C.R. Biologies, 330, 2007, pp. 189-198
[24] The surface area of French Guiana is 84,000 km2 (close to that of Portugal), 75,000 km2 of which is forest (about 50% of metropolitan forests).
[25] Gourlé-Fleury S., Guehl J.M. & Laroussinie O. Ecology and Management of a Neotropical Forest. Lessons drawn from Paracou, a long-term experimental research site in French Guiana. Elsevier, 311p, 2004.
[26] Saint-Jean D. & Pellet E. Explorateurs d’Amazonie. Aventuriers de la Science en Guyane. Ibis Rouge Éditions, 2008 (Preface Alain Pavé). (in french)
[28] One must be very careful in the use of words, initially having an anthropic meaning: there is no intention behind competition and cooperation in ecological systems. These mechanisms are set up spontaneously.
[29] Norten N., Angarita H. A., Bongers F., Martinez-Ramos M., Granzow-de la Cerda I., van Breugel M., Lebrija-Tejos E., Meave J.A., Vandermeer J., Bruce Williamson G., Finegan B., Mesquita R. & Chazdon R.L. Successional dynamics in neotropical forests are as uncertain as they are predictable. Proc. Nat. Acad. Sci. USA, 112, 8013-8018, 2015.
[30] Anna Roosevelt is one of the most famous archaeologists and ecologists working on the Amazon. It extends a family interest in this region from her great-grandfather Theodore Roosevelt, who was President of the United States (1901-1909) and stayed many times in the Amazon after his two terms. Among Anna Roosevelt’s many publications, we can mention: Roosevelt A.. Twelve Thousand Years of Human-Environment Interaction in the Amazon Floodplain. Advances in Economic Botany, Vol. 13, New York Botanical Garden. pp. 371-392, 1999.She was a member of the Scientific Committee of the Amazon Programme of the CNRS.
[31] Reich D. et al. Reconstructing Native American population history. Nature, 488, 370-375, 2012. This work was supported by the CNRS Amazon Programme.
[32] Rostain S., Amazonie : les 12 travaux des civilisations précolombiennes. Belin, Paris, 2017; & Rostain S., Amazonie un jardin sauvage ou une force domestiquée. Essai d’écologie historique.. Actes Sud, wandering, 2011.
[33] Molino J.F., Mestre M. & Odonne G. La biodiversité de l’Amazonie, un héritage des Précolombiens ? Research, 527, 67-71, 2017.
[34] Jérémie S., Dambrine E.. Impact des occupations amérindiennes anciennes sur les propriétés des sols et la diversité des forêts guyanaises. In, Alain Pavé and Gaëlle Fornet, Op. Cit. (in French)
[35] McKey D., Rostain S., Iriarte J., Glaser B., Birk J.J., Holst I. &Renard D., “Pre-Columbian Agricultural Landscapes, Ecosystem Engineers, and Self-organized Patchiness in Amazonia”, Proc. Nat. Acad. Sci. USA, 107, 2010, p. 7823-7828. CNRS press release, March 13, 2010.
[36] The International Centre for Tropical Agricultural Research in Cali, Colombia, maintains a living collection of about 5000 varieties. These collections are one of the ways to maintain biodiversity.
[39] Renca (Reserva Nacional do Cobre e Associados) was created in the early 1980s for future mining. In the absence of this exploitation it has become a de facto nature reserve. This status was threatened in 2017 by then Brazilian President Michel Temer. In the face of national and international protest, the status was maintained.
[40] Raby M. The Colonial Origin of Tropical Field Station. Am. Scientist, 105, 216-223, 2017.
[41] Legay J.M. & Barbault R. (Dir.). La révolution technologique en écologie. Masson, 1995. (in French)
[42] Pavé A., Comprendre la biodiversité, vrais problèmes et idées fausses. Threshold Edition, 2019. (in French)
[43] Worster D. The Wealth of Nature, Environmental History and the Ecological Imagination. Oxford University Press, 1993.
[44] These two pre-Socratic philosophers lived well at the same time, but were very distant from each other. Elée, in the south of present-day Italy, for Parmenides, and Ephesus, on the west coast of this Turkey, for Heraclitus. This debate, undoubtedly virtual, has been reconstructed from the writings of these two philosophers and those who were inspired by them. Broadly speaking and in current terms, Parmenides defended the idea of a stationary world, with observed movements and changes responding to permanent mechanics. Heraclitus had a conception of a world in constant evolution (“you never bathe twice in the same river“), with often chaotic dynamics. This debate has implicitly marked the history of ideas, it is even still very present, especially in ecology (see, the previous reference).
[45] Casseta E & Delors J (Eds). La biodiversité en question. Enjeux philosophiques, éthiques et scientifiques. Editions Matériologiques, 2014, Paris. (in French)
[46] Multi-agent technology has already proven its effectiveness, particularly in modelling socio-ecosystems. The concept of accompanying modelling seems well suited to this type of situation. Ref. : Collectif Comod, La modélisation comme outil d’accompagnement, Natures Sciences Sociétés, 13, 165-168, 2005 (in French). It is not out of place to also draw inspiration from the work of automation engineers, who are curiously little known outside the field of industrial engineering. In the 1980s, in Rhône-Alpes, the cooperation initiated between biometricians in Lyon and scientists specialists of automatic control, in Grenoble, extended within the Edora club of Inria, was very effective, particularly in establishing this cooperation.