Deforestation, fragmentation and pollution of habitats, intensification of agriculture and livestock farming, urbanization, massive emissions of greenhouse gases into the atmosphere, acidification of the oceans, intensive industrial fishing, transportation of species… Our expanding species does exert increasing pressures on the planet’s ecosystems, through its many activities. How do species and ecological communities react to these pressures? Can we predict their dynamics? Given the multiplicity of species and of their interactions, the first step here will be to identify the main drivers of the adaptation of species and communities to changes in their physical and biological environment. Then, we will take these mechanisms into account to understand and possibly anticipate current dynamics, in this period of global changes linked to the intensification of human activities.
Since Darwin, we have known that species change over time as a result of natural selection, which sieves the fittest individuals within the populations, or more precisely those who better transmit their biological characteristics (with their genes) to new generations. This process of evolution and adaptation allows species to respond to the usual variations in their living conditions, on different time scales.
Research in ecology and evolution has confirmed for the past sixty years that species are adapted to the variability of the living conditions usually encountered by individuals, on the one hand, but also to the relative frequency of “catastrophic” events [1] encountered by the populations. Let us see how.
1.1. Niche width and ‘phenotypic’ plasticity
According to Hutchinson [2], any species or population may be characterized by its “ecological niche”, which may be defined as the “place” occupied by this species in an ecosystem, i.e. by the set of its physical, chemical and biological requirements. As products of natural selection, these ecological requirements evolve over generations, with the physiology and behaviour of the species. Thus, the “width” of the ecological niche of each species is adapted to the variations in living conditions usually encountered by individuals, in time and space (see Focus From niche construction to the destruction of ecosystems).
Figure 1. Populating estuaries, lagoons and Mediterranean coastlines, jumping mullets (Liza saliens) tolerate wide fluctuations of water salinity. [Source: Ray eye, CC BY-SA 2.5, via Wikimedia Commons]For example, pelagic (i.e. open ocean) fish and invertebrate species are often stenohaline because the salinity of the water off the coast varies very little (around 32 g/l); estuarine species, on the other hand, are necessarily euryhaline, as they are adapted to wide fluctuations in salinity.
Thus, natural selection prepares the individuals for the usual variations in their living conditions, whether long-lasting or recurrent, by favouring morphological, physiological and/or behavioural modifications adapted to these variations. Individuals then show a certain adaptability, or so-called “phenotypic” plasticity, which can be implemented during their development (and therefore be irreversible), or continue throughout life, accompanying or preceding environmental variations.
Among the irreversible phenotypic adaptations, let us quote the formation of a protective helmet in daphnia, conditioned by the presence of predatory fish in their initial habitat [3]; or the caste determinism in social insects (social bees, ants, termites, ..), whose larvae become workers, soldiers (in termites) or reproducers according to the diet given to them by the workers of the colony.
When environmental variations are recurrent or cyclical, the plasticity of individuals can be reversible. Thus, the phenology of species is set to the rhythm of daily (nycthemeral cycle) and seasonal variations of their habitats. In temperate regions, deciduous trees are sensitive to day length, or photoperiod. Each autumn, they shed their leaves, produce buds and go dormant for the winter.
Figure 2. As an adaptation to the annual cyclic changes of their environment, alpine marmots (Marmota marmota) build up ample fat reserves in late summer, before lining their burrows with dry grass and locking themselves in as a family for the winter. [Source: David Monniaux, CC BY-SA 3.0, via Wikimedia Commons]They accompany many animal species, especially insects and terrestrial vertebrates (mammals, reptiles and amphibians, but not birds) that take refuge in a shelter to face the winter in a state of torpor (hibernation, winter lethargy, diapause…). Among birds, warm-blooded organisms incapable of hypothermic change, some species such as jays, nuthatches and nutcrackers stock up on seeds at the end of the summer, which will feed them in winter, while others (insectivores in particular, such as swifts and swallows) stock up on fat and migrate to milder latitudes, before coming back to nest in the summer.
Similarly, the 24-hour activity cycle (circadian cycle) of all vertebrates, punctuated by the secretion cycle of a brain hormone, melatonin, is an adaptation to the length of the days – and the alternation of day and night – on our planet.
Figure 3. Tobacco plant. When its leaves are attacked by a predator, like many other plants, the tobacco plant defends itself by synthesizing phenolic compounds that are toxic to herbivores. In addition, like Alder, Wormwood or Sugar Maple, it emits an alarm pheromone (volatile compound) that induces the same synthesis in nearby plants of the same species. [Source: Forest & Kim Starr, CC BY 3.0, via Wikimedia Commons]However, organisms are not simple automatons, programmed to feed, defend, and reproduce in an environment of predictable and controlled variations: most are also capable of responding to unexpected environmental variations with appropriate behavior, revealing a certain behavioral elasticity.
Plants are not to be outdone. Plants can react to daily, not only cyclical (light intensity, temperature…) but also unpredictable (predation, infection, competition…) variations of their local environment by adapted behaviors. We may observe, for example, the orientation and/or opening of flowers according to their exposure to light, the synthesis of toxic anti-predator compounds, the emission of attractive pheromones for allied species or alarm pheromones intended for neighbouring plants of the same species, in case of aggression.
Figure 4. Common octopuses are known for their propensity to explore and ‘manipulate’ new objects, their excellent visual memory and their learning abilities. [Source: Albert Kok at Dutch Wikipedia (Original text: Albert Kok), Public domain, via Wikimedia Commons]While it exists in plants, which have no real sense organs, behavioral plasticity is a general property of animals. It increases with their learning capacities (cognitive and practical abilities), as well as with their sociality (complexity of social groups) and mobility. It has been observed in some social invertebrates, such as the Honey Bee, and in non-social invertebrates, such as the common octopus, and peaks in mammals and birds, which have high cognitive capacities, high energy requirements and often complex social systems [4].
Physiological tolerance, morphological or physiological adjustments and behavioural flexibility are three types of mechanisms that allow individuals to respond adaptively to habitual variations in their environment. These mechanisms contribute to the adaptive potential of species to changes in their habitats and living conditions, whether these are caused by other “actors”-humans, in the case of most current environmental changes – or by their own activities (see Focus on From niche construction to the destruction of ecosystems).
1.2. Habitat stability and demogenetic strategies
On a larger time scale, covering several generations, the biological characteristics of species can also evolve in response to fluctuations in the environment that cannot be predicted at the individual level.
The mechanisms of long-term adaptation of species to variations in their habitats are not only demographic but also genetic, since they rely on the diversity and genetic evolution of populations. Research in evolutionary ecology has confirmed since the pioneering work of Mac Arthur and Wilson [5] in this field that the “demogenetic strategies” (both demographic and genetic) of species are adapted to the relative frequency of catastrophic-type events, inflicting high mortality on populations.
one, based on “quantity“, of rapid multiplication of individuals (known as “r“, for reproduction) and random dispersal, in temporary habitats that are unpredictable at individual scale
the other, based on “quality“, with late and limited but competitive reproduction (known as “K“), in more predictable and saturated habitats.
According to Pianka [6], the main life traits favored by r-type selection are:
reproduction at an early age, associated with a short lifespan and smallsize,
high fecundity,
early dispersal of offspring (seeds, eggs, larvae…), by passive or active mobility.
And the life traits associated with the “K” strategy are:
late reproduction, correlated with a long life span and large size,
Furthermore, the genetic diversity of species increases with the number of individuals; thus, it increases with the area of favorable habitats, with their connectivity, and with the intrinsic growth rate of populations.
The ‘r’ strategy, in response to episodic decimation
Figure 6. Colonization of nutrient-rich nectarines by microscopic fungi known as ‘moulds’. [Source: Roger McLassus 1951 assumed (based on copyright claims), CC BY-SA 3.0, via Wikimedia Commons]This is the usual strategy of microorganisms – such as bacteria, protists, fungi… and viruses! – inhabiting temporary habitats (animal or plant hosts, seasonal habitats, organic waste…), as well as of macroscopic algae and fungi, able to quickly colonize any new favorable habitat.
Taking advantage of the fecundity and mobility of individuals, rather than of their adaptations and skills in terms of local survival, the “r” strategy (of rapid multiplication and dispersal) allows small prolific species to rebound after local decimation. Random migration and genetic mutation accelerate their genetic evolution and adaptation to the diversity (and unpredictable spatial distribution) of their potential habitats. The “random dispersal” of large numbers of close related individuals following the “r” strategy favours the discovery and exploitation of new resources (e.g. a new host species) by some of them, and thus the adaptation to new living conditions. These characteristics increase the probability of long-term survival of species (metapopulations) adapted to unstable habitats[10].
In plants, this ‘r’ strategy is used by many annuals and grasses, which disperse pollen and seeds in large quantities over good distances (via wind or animal transport) and rapidly colonise open habitats before being browsed, trampled, burnt, wilted or replaced by other, more competitive and long-lived species.
This capacity for resilience and rapid evolution of populations, which increases with their intrinsic growth rate and genetic diversity, seems to be a strength of microorganisms and other prolific species in the face of current global changes. This is how bacterial strains resistant to certain antibiotics, fungal lines resistant to antifungals, herbicide-resistant messicolous plants and insecticide-resistant “pest” insects appear from time to time and then flourish (see Focus on The demogenetic strategy of bees).
The ‘K’ strategy, poorly adapted to the disruption of habitats
Figure 8. Female hippo and her calf. Like most large ungulates (antelopes, bighorn sheep, horses, rhinoceroses, ..), female amphibian hippos reach sexual maturity between 4 and 7 years of age and give birth to a single calf after a long gestation period (8 months) – at a rate of one calf every two or three years in their case. [Source: Jnissa, CC BY 2.0, via Wikimedia Commons]In contrast to the ‘r’ strategy, the demogenetic (‘K’) strategy is that of long-lived, low-fertility mammals, such as ungulates, marine mammals, and great apes, which raise at best one calf per year in relatively small populations (low genetic diversity). This strategy can penalise them in the face of upheavals in their living conditions (see Focus on Adaptations to climate change on the dynamics of mammals). The survival of populations then depends on the intensity of local and regional upheavals, as well as on the physiological and behavioural plasticity of individuals.
It should be noted, however, that the fecundity of sexual species varies, among other factors, with the capacity of females to invest time and energy in the survival and reproduction of each of their offspring. In the absence of parental care, natural selection favours the most fertile females.
Thus, in plants with limited parental capacities, the strategy of large, long-lived species such as trees, which are adapted to the stable and saturated habitats of forests, is accompanied by good fecundity that increases with the size and age of the individuals. Similarly, with little capacity to protect their eggs, which are dispersed at sea, the females of large pelagic fish species such as tuna and cod invest in numbers (high fecundity) and not in the individual survival of their offspring.
In general, the complexity of the evolutionary strategies of species increases with the diversity of their interactions. For example, despite their small size and the high fecundity of the (few) reproductive females, eusocial species such as honeybees, termites and naked mole rats are more ‘K’ than ‘r’ strategists, since they restrict their reproduction (to a small fraction of the colony members) and rely on collective young rearing and intragroup cooperation to increase the predictability of their environment, and thus the permanence of their colonies (see Focus on Adaptations to habitat anthropization).
2. Adaptation potential and global change
We may distinguish three main factors of species adaptation to environmental changes:
the genetic diversity of populations, a reservoir of possible responses to unpredictable changes in the environment,
their “intrinsic” growth rate, which favours their resilience after decimation, and
the phenotypic plasticity (morphological, physiological and behavioural) of individuals.
While prolific and small species (type ‘r’), in all systematic groups, often combine the first two factors, the third characterizes animal species with high cognitive capacities, such as mammals and birds. However, these are partly ‘K’ species of large size and low fecundity, with small numbers, poorly equipped on the ‘demogenetic’ level to cope with large and rapid changes in their environment. Under these conditions, can we predict the dynamics of species and communities in response to current global changes, which can be characterized by their diversity, ubiquity and speed?
2.1. What are the main species responses?
Figure 9. Deer at the edge of a wood. Gleaning its food in forests as well as in fields and meadows, taking advantage of the absence or rarity of wolves and other large predators, the roe deer is a generalist species in expansion in Europe. [Source: Sylvouille at French Wikipedia, CC BY-SA 1.0, via Wikimedia Commons]Depending on the biological and demographic characteristics that underpin the species’ potential for adaptation, one may anticipate and verify in the field:
the expansion of generalist species (‘r’ and ‘K’, including our own!), in all ecosystems modified by human activities – the case of wild boar, roe deer, great tit, woodcock and crow in Europe; the case of olive baboons in central Africa… ;
the resilience of prolific ‘r’-type species (with high fecundity, early reproduction, high mobility of juveniles or adults), after local or regional decimation – illustrated for instance by the episodic infestations of migratory locusts, corn borers, ground voles and other so-called ‘crop pests’ in intensive monoculture regions;
the adaptation and rapid genetic evolution, or even speciation, of ‘r‘ species confronted with new living conditions, in modified ecosystems – the case of new antibiotic-resistant bacterial lines and recombinant viruses involved in infectious disease epidemics, which have been on the increase for several decades [11]; the case of pesticide-resistant insects and annual plants (see Focus on Adaptations to habitat anthropization) [12] ;
the slower behavioural adaptation and genetic evolution of generalist ‘K’ species, commensals of humans, expanding in these modified ecosystems – this is expected for the expanding ‘K’ species mentioned above: deer in Europe, savannah baboons in Africa… ;
the more or less rapid displacement of ‘r’ and ‘K’ species to more favourable living conditions in response to global warming – for example, the displacement of bark beetles (xylophagous beetles), leafrollers (phytophagous caterpillars) and other so-called ‘forest pests’ towards the poles and at higher altitudes.
But also:
the decline and disappearance ofmany specialist ‘K’ species, weakened by the fragmentation and/or progressive transformation of their habitat and unable to rebound after local decimation (see the focus on Mammals and global change, on the dynamics of mammals),
the rarefaction and disappearance of a fraction of ‘r’ species, mainly specialists, confronted with the disappearance or radical transformation of their habitats and resources (e.g. invertebrates and fish in coral reefs, insects in regions of intensive agriculture);
2.2. What are the dynamics of communities and ecological networks?
In all ecosystems modified by human activities, the expansion of generalist species at the expense of specialist species should result in a homogenisation of regional faunas and floras [14], [15] and a decline in the total number of species on a global scale (unless there is sufficient diversification through speciation in isolated emerging habitats, cf. [16]).
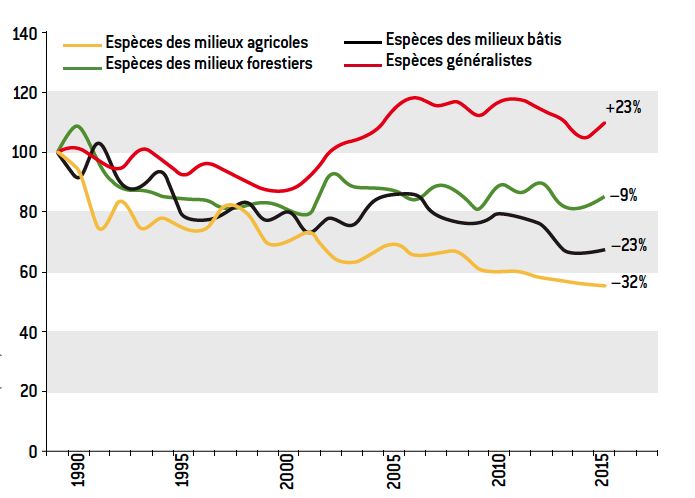
Figure 12. Temporal Monitoring of Common Birds (STOC), since 1989: STOC indicator by habitat type. While generalist species are expanding in France (and elsewhere), species specialising in agricultural and other habitat types are declining. As a result, the specialization index of local bird communities has been declining over the years, for several decades. [Source: CRBPO, Vigie-Nature, MNHN]Thus in Europe, in response to the intensification of agriculture, generalist bird species (chickadees, starlings, carrion crows, wood pigeons…), which frequent fields as well as towns, villages and/or forests are in expansion in (degraded) fields for twenty years. Conversely, birds specialist of agricultural environments (e.g., linnets, hoopoes, grey partridge, crested lapwings, etc.) are rarifying (see fig.12). To monitor this dynamic in bird community composition, researchers have developed a community specialisation index, which increases with habitat disturbance [17] and confirms the homogenization of France’s bird populations over the past 20 years [14] and [18].
Ecosystems are complex organized systems of limited resistance and resilience, formed by networks of co-adapted species, evolving in interaction with each other and with their physical environment for many generations. The rapid change in community composition, coupled with the different rates of adaptation of species to changes in their environment (among which climate changes), are desynchronizing and disorganizing these networks. These changes increase the risks of ecosystems switching to another steady state, unfavorable to many species often including humans [19], [20] (Figure 13).
Figure 14. Pullulation of common jellyfish (Aurelia aurita) on a Danish coast. [Source: Photo Vandmaend, CC BY 3.0, via Wikimedia Commons]Over time, pressures on ecological communities and networks increase as human societies take over land and “resources”. In fact, the conversion or degradation of habitats rich in biodiversity (such as forests, marshes, fish-filled rivers and seas…) into impoverished habitats (cultivated fields, polluted rivers, “overfished” seas), with less biotic capacity, reduces the total amount of resources available for wildlife by at least a quarter [21]. This restriction implies not only a reduction in the abundance and biomass of the animal and plant communities concerned [22], [23], but also in the number of species [24]. It thus favours a shortening offood webs[25], [26] with an increased risk of ecosystem collapse (see focus From niche construction to the destruction of ecosystems) and a shift to another stable state unfavourable to many species including our own [27], [28], [29].
3. Messages to remember
Through its many activities and interactions, each species modifies its environment and builds its ecological niche over generations.
Some species, known as “environmental engineers“, have a strong impact on their physical and biological habitats, and thus on the structure and functioning of local communities. One of them – ours! – has distinguished itself in recent centuries by the diversity and increasing scale of its impacts on terrestrial and aquatic ecosystems, which are currently undergoing global change.
The response of each species to changes in its physical and biological environment depends on external factors, such as the magnitude and frequency of these changes, and internal factors, such as its behavioural plasticity and reproductive fecundity.
Species are adapted to the variability of living conditions to which individuals are usually exposed, over many generations. Their ecological niche is therefore more or less wide or narrow, reflecting these variations: they are more or less generalists or specialists.
In addition, they are adapted to the frequency ofextreme events, which decimate populations, by their demographic dynamics.
We can thus distinguish species according to their “demogenetic strategy“, adapted or not to the extent and speed of the global changes underway.
Ecosystems and ecological networks, made up of numerous species interacting with each other and with their habitat, are complex, organized and adaptive systems with limited resilience.
Beyond a certain threshold of disturbance, ecosystems switch to another state, characterized by a different structure (species networks) and a different operating regime.
This is currently the case for many ecosystems faced with global changes. These ecosystem chain shifts are not only detrimental tomanyspecies but also to human societies, which are victims of their increasing pressure on ecosystems and the biosphere.
This article is a revised, adapted and updated version of the article “Quelles réponses des espèces et communautés écologiques aux changements globaux?” (R80a) by the same author, dated June 2018, online on the interactive platform of the Société Française d’Ecologie et Evolution (SFE2, https://www.sfecologie.org/regards/). (Regard edited by Sébastien Barot, SFE2, in February 2018).
Notes and references
Cover image. Pullulation of common jellyfish (Aurelia aurita) on a Danish coast [Source: Photo Vandmaend, CC BY 3.0, via Wikimedia Commons]
[1] Catastrophic event, for a population: inflicting high mortality.
[4] Teyssèdre A., 2006. Les clés de la Communication animale. Delachaux et Niestlé, Paris.
[5] MacArthur R.H. & E.O. Wilson, 1967. The theory of island biogeography. Princeton Univ. Press, Princeton, NJ.
[6] Pianka E.R., 1970. On r and K selection. Am. Nat. 104:592-597.
[7] Reznick D., M.J. Bryant & F. Bashey , 2002. R- and K-section revisited: The role of population regulation in life history evolution. Ecology 83(6):1509-1520.
[8] Beaumont H.J.E., Gallie J. et al. 2009. Experimental evolution of bet hedging. Nature 462:90-92.
[9] Botero C.A., F.J. Weissing, J. Wright & D.R. Rubenstein, 2015. Evolutionary tipping points in the capacity to adapt to environmental change. Proc. Natl. Acad. Sci. USA 112:184-189.
[10] Starrfelt and Kokko 2012. Bet hedging: a triple trade-off between means, variances and correlations. Biol. Rev. 87, pp. 742-755.
[13] Teyssèdre A., 2018. Les mammifères face aux changements globaux. Regards et débats sur la biodiversité, SFE, regard R80b, June 2018.
[14] McKinney M.L. & J.L. Lockwood, 1999. Biotic homogenization: a few winners replacing many losers in the next mass extinction. T.R.E.E . 14:450-453
[15] Clavel J., Julliard R. and V. Devictor, 2010. Worldwide decline of specialist species: toward a global functional homogenization? Front. Ecol. Environ. 2:222-228.
[16] Thomas C.D., 2015. Rapid acceleration of plant speciation during the Anthropocene. Trends Ecol. Evol. 30:448-455.
[17] Julliard R., J. Clavel et al. 2006. Spatial segregation of bird specialists and generalists in bird communities. Ecol. Letters 9:1237-1244.
[18] Clavel J., 2011. L’Homogénéisation biotique. Regards et débats sur la biodiversité, SFE, regard n°16, April 2011.
[19] Folke C., S. Carpenter et al. 2004. Regime shifts, resilience and biodiversity in ecosystem management. Ann. Rev. Ecol. Syst. 35:557-581.
[20] Cardinale B. et al. 2012. Biodiversity loss and its impact on humanity. Nature 486:59-67.
[21] Haberl H., 2007. Quantifying and mapping the human appropriationof net primary production in earth’s terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 104:12944-12947.
[22] Gaston K.J., T.M. Blackburn, K. Klein Goldewijk, 2003. Habitat conversion and global avian biodiversity loss, Proc. R. Soc. Lond. B 270:1293–1300.
[23] Smil V., 2011. Harvesting the Biosphere: the Human Impact. Pop. Dev. Rev. 37(4):613-636.
[24] Teyssèdre A. & D. Couvet, 2007. Expected impact of agriculture expansion on the world avifauna. C. R. Acad. Sci. Biol. 330:247-254.
[25] Pauly D, V. Christensen V, J. Dalsgaard J, R. Froese & F.S.B. Torres, 1998. Fishing down marine food webs. Science 279:860-863.
[26] Watson R. & D. Pauly, 2001. Systematic distortions in world fisheries catch trends. Nature 414(6863):534-536.
[27] Folke C., S. Carpenter et al. 2004. Regime shifts, resilience and biodiversity in ecosystem management. Ann. Rev. Ecol. Syst. 35:557-581.
[28] Cardinale B. et al. 2012. Biodiversity loss and its impact on humanity. Nature 486:59-67.
The Encyclopedia of the Environment by the Association des Encyclopédies de l'Environnement et de l'Énergie (www.a3e.fr), contractually linked to the University of Grenoble Alpes and Grenoble INP, and sponsored by the French Academy of Sciences.
The articles in the Encyclopedia of the Environment are made available under the terms of the Creative Commons BY-NC-SA license, which authorizes reproduction subject to: citing the source, not making commercial use of them, sharing identical initial conditions, reproducing at each reuse or distribution the mention of this Creative Commons BY-NC-SA license.
Since the Spanish flu of 1918, the term pandemic has become commonplace. The last infectious…
Emmanuel DROUET, Doctor of Pharmacy, Professor at the Université Grenoble-Alpes, Professor at Institut de biologie structurale UMR 5075 CNRS UGA CEA, Grenoble.