


Le système racinaire des plantes : de l’ombre à la lumière
PDF
Les racines représentent une cible de choix pour l’amélioration des plantes cultivées pour accompagner la transition vers une agriculture plus frugale et respectueuse de l’environnement. Pourtant, les caractères racinaires ont longtemps été négligés dans les programmes d’amélioration, notamment car la première révolution verte a été basée sur l’utilisation massive d’eau et d’engrais. Les changements globaux incitent désormais à une utilisation plus raisonnée des intrants agricoles afin de réduire la dégradation des sols, la pollution des eaux et de l’air, et l’épuisement des ressources. Répondre aux besoins de production tout en limitant l’utilisation d’eau et d’éléments minéraux est un défi majeur. Quel rôle peuvent jouer les racines, organes nourriciers de la plante, dans le développement d’une agriculture plus durable et résiliente face aux climats actuels et futurs ? Un rôle essentiel, comme le montrent les recherches qui permettent de mieux comprendre leur développement, leurs fonctions, leur diversité et leurs adaptations à l’environnement, les faisant passer de l’ombre à la lumière.
1. Les racines prélèvent l’eau et les nutriments pour la plante
Les racines sont les organes des plantes supérieures spécialisés dans l’exploration du sol et le prélèvement de l’eau et des ions minéraux (Lire Nourrir les plantes en polluant moins ?). Elles jouent un rôle primordial pour leur survie et leur développement. Chaque plante possède plusieurs catégories de racines organisées en un système racinaire. Cet ensemble de racines ancre la plante dans le sol et communique avec la ou les tiges, qui portent les feuilles, les fleurs et les fruits. Le système racinaire se développe par la formation répétée de nouvelles racines et leur croissance en longueur. La croissance et la formation des racines dépendent étroitement de l’environnement dans lequel elles se développent. Les racines permettent ainsi à la plante de s’adapter, malgré son immobilité, au caractère hétérogène et variable des ressources et des contraintes de son milieu de vie (Lire La vie fixée des plantes et ses contraintes, La quête inlassable de l’eau par les plantes & Besoins en eau des plantes : comment les satisfaire ?).
1.1. Architecture et anatomie du système racinaire

Deux grands types de systèmes racinaires peuvent cependant être distingués [1]. Le système racinaire « pivotant », retrouvé chez les Angiospermes dicotylédones (comme le colza ou le pois) et des Gymnospermes (telles que le pin), se caractérise le plus souvent par la formation de racines latérales sur une racine séminale principale qui croît dans le sol telle un pivot (Figure 1a) [2]. La majorité des Angiospermes monocotylédones (comme le blé ou le riz) possèdent quant à elle un système racinaire de type « fasciculé » qui se caractérise par un bouquet de plusieurs racines séminales et surtout de nombreuses racines dites coronaires relativement équivalentes en taille et se formant à partir de la base des tiges (Figure 1b). Ces racines portent elles-mêmes des racines latérales. Certaines plantes possédant un système racinaire de type « fasciculé » peuvent développer plus d’une centaine de racines coronaires.
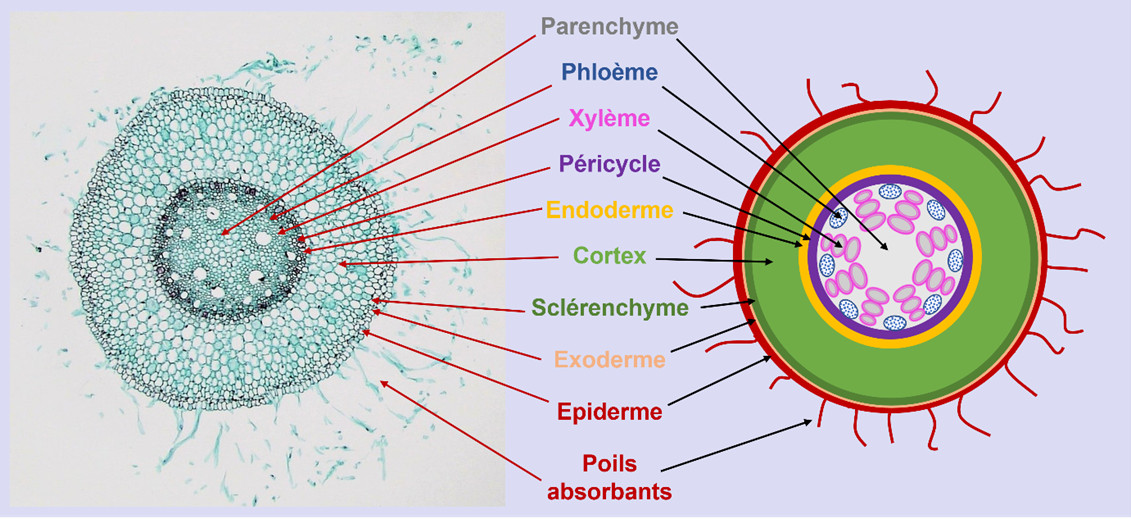
Ces différents tissus racinaires possèdent des propriétés qui permettent à la racine de moduler ses capacités d’échanges avec le sol :
- Les poils absorbants permettent notamment d’augmenter drastiquement la surface d’échange de la racine pour faciliter l’acquisition de l’eau et des nutriments.
- Des barrières pariétales plus ou moins imperméabilisées entourant les cellules de l’endoderme et du cortex externe régulent le passage de l’eau et des nutriments vers les vaisseaux conducteurs [3].
Les tissus vasculaires sont eux spécialisés dans les échanges entre les racines et les organes aériens :
- Les vaisseaux du xylème contiennent la sève brute composée d’eau et d’éléments minéraux qui circule des racines vers les organes aériens.
- Les vaisseaux du phloème contiennent la sève élaborée riche en produits de la photosynthèse qui circule des organes aériens vers les racines.
Chez les espèces pérennes, les racines âgées s’élargissent au fil des années par l’ajout de nouvelles couches de tissus. Ces tissus dits secondaires forment, entre autres, du liège en périphérie et du bois au centre de la racine.
1.2. Développement et plasticité du système racinaire
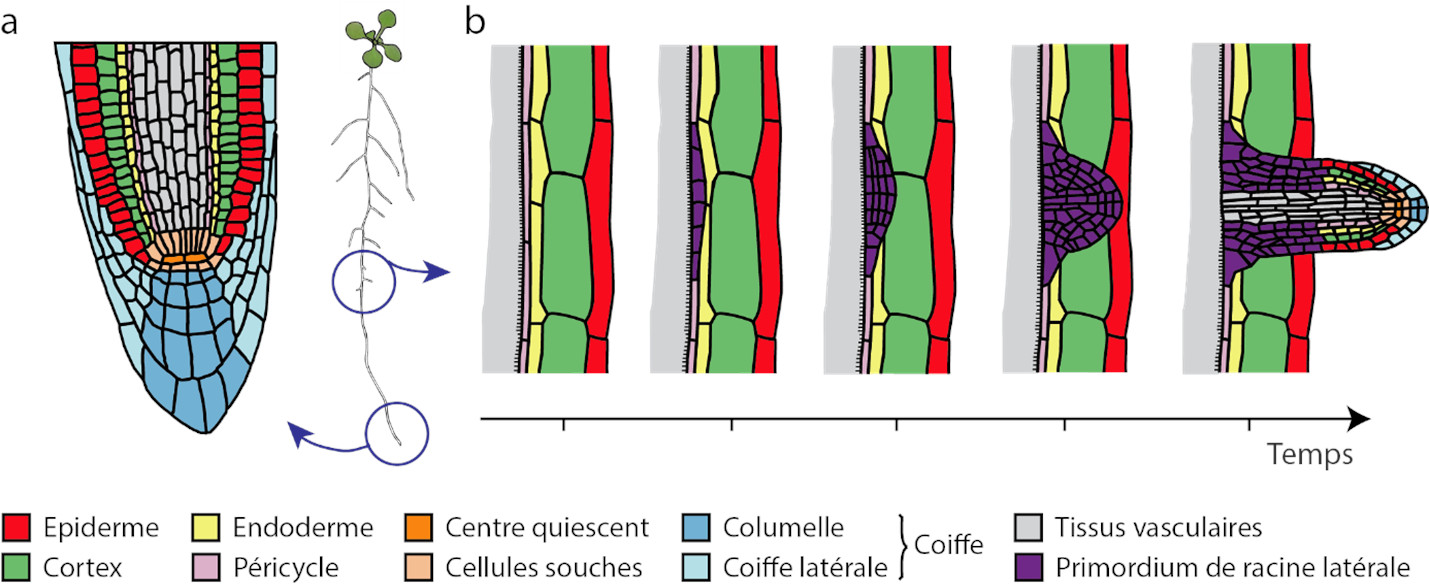
Le développement des racines dans le sol est très sensible à l’environnement. La croissance, le développement et les propriétés fonctionnelles des racines sont influencées par des signaux circulant dans la plante et par la perception d’informations en provenance du sol. Ces modifications du système racinaire en réponse aux conditions changeantes de l’environnement sont cruciales pour l’adaptation de la plante et sont désignées par le terme de plasticité racinaire.
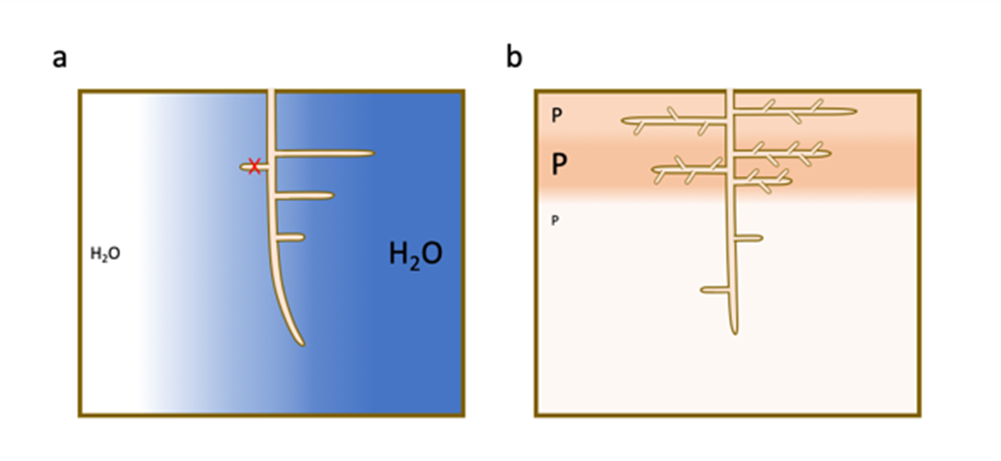
- l’auxine s’accumule préférentiellement dans les zones de la racine en contact avec le sol humide et y stimule la formation d’un nouveau méristème racinaire.
- l’acide abscissique, hormone de réponse au stress hydrique, s’accumule préférentiellement dans les zones sèches du sol et y réprime la formation des racines latérales.
Comme pour l’eau, une forte accumulation locale de phosphore et de nitrate dans des sols généralement pauvres induit une prolifération préférentielle de racines latérales dans ces zones [7] (Figure 4b).
Ainsi, le système racinaire de la plante est une structure complexe se développant de façon continue dans le sol en s’y adaptant de façon dynamique. Les caractères adaptatifs de plasticité racinaire sont favorables à la nutrition de la plante dans des conditions où les ressources sont limitées. De plus, par leur activité, ces racines interagissent avec le sol et modifient ses caractéristiques.
2. Les racines interagissent avec le sol et les communautés microbiennes qui s’y trouvent
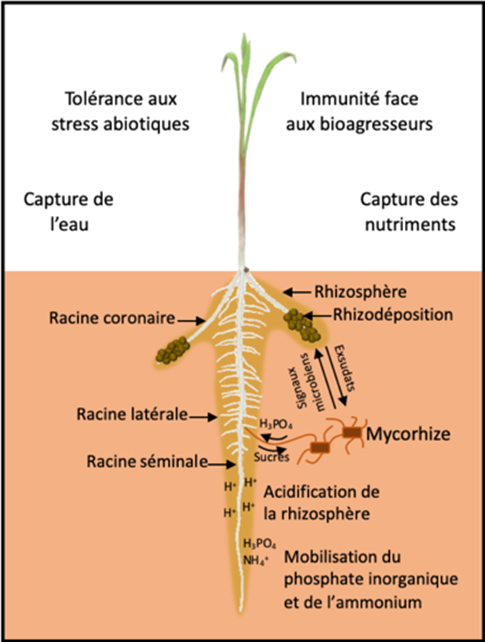
2.1. Influence des racines sur les caractéristiques du sol
Certains nutriments sont présents dans le sol sous une forme chimique inaccessible pour les plantes. L’exsudation racinaire contribue à rendre ces éléments plus mobilisables et absorbables par les racines. Des mécanismes d’acidification du sol par l’exsudation de protons ou de phytosidérophores [9] favorisent ainsi l’acquisition du fer chez certaines plantes [10]. L’exsudation racinaire est également un processus plastique et dépend de l’environnement dans lequel se développe la racine. Des plantes de lupin blanc se développant dans un sol pauvre en phosphore produisent, par exemple, des clusters de racines courtes et denses appelés « racines protéoïdes » [11]. Ces racines sécrètent de grandes quantités de molécules acides capables d’abaisser le pH du sol, rendant les ions phosphore plus assimilables. Chez cette espèce végétale adaptée aux sols pauvres en nutriments, cette plasticité de l’activité des racines est un mécanisme d’adaptation majeur. Une expérience a pu montrer qu’une plante de lupin se développant sur un sol pauvre en phosphore peut sécréter jusqu’à 25 fois plus d’exsudats qu’une plante se développant sur un milieu riche en phosphore [12].
L’exsudation, contribue également à la formation d’un manchon racinaire. Ce manchon correspond à l’agrégation, autour des racines, d’un sol riche en mucilage et autres molécules carbonées. Il participe chez certaines espèces à améliorer l’acquisition de l’eau. Chez le pois chiche, un manchon racinaire important a ainsi été associé à une meilleure capacité de la plante à tolérer le stress hydrique [13]. Cette agrégation des particules autour des racines pourrait favoriser l’humidité du sol au contact de l’épiderme racinaire et contribuer à maintenir la zone de contact et d’échange entre le sol et la racine en conditions de sécheresse. De plus, ce sol agrégé riche en exsudats représente une source de carbone importante pour les microorganismes susceptibles d’interagir de façon bénéfique avec la plante.
2.2. Mobilisation des microorganismes par les racines
La rhizosphère est généralement microbiologiquement très active. Certains exsudats racinaires agissant comme des signaux attractifs pour des microorganismes [14]. Des molécules de strigolactone [15] sécrétées par la plante dans la rhizosphère stimulent ainsi la germination des spores de champignons mycorhiziens à arbuscule. Les molécules produites par le champignon favorisent en retour le déclenchement de processus racinaires qui permettent l’établissement d’une symbiose entre la plante et le champignon (Figure 6). Au cours de cette symbiose, l’exploration d’un volume de sol très important par les hyphes mycéliens du champignon contribuent à l’approvisionnement de la plante en eau, en phosphore et en d’autres éléments minéraux. En retour, le champignon bénéficie des molécules carbonées produites par la photosynthèse de la plante.
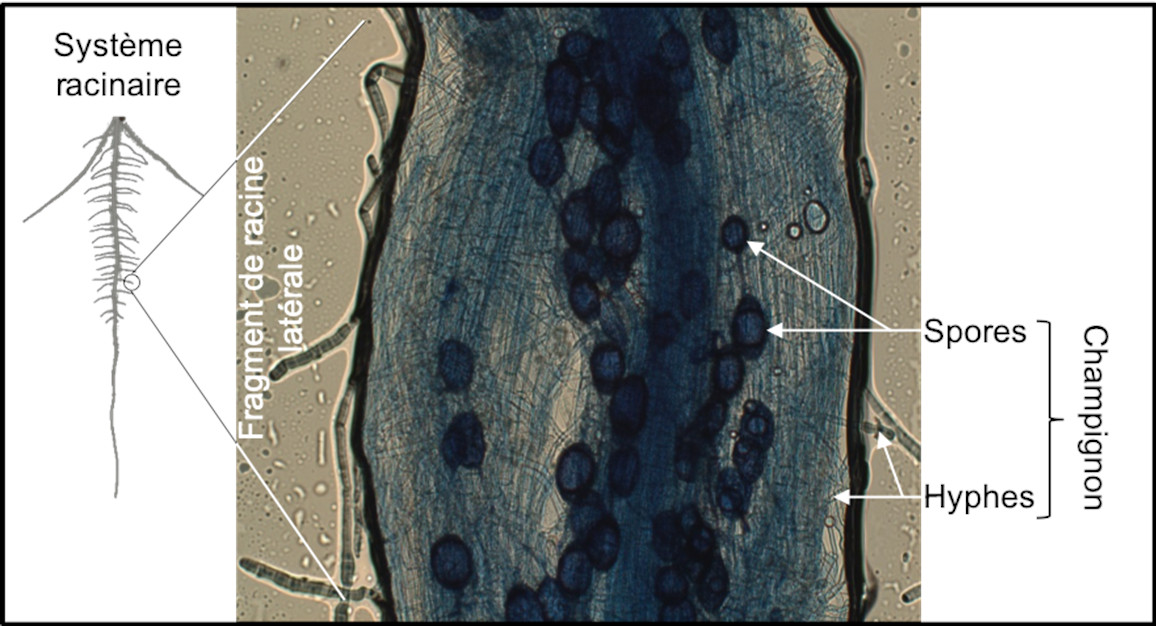

Par leur développement continu et plastique, leur propriétés adaptables et leur actions bénéfiques sur la rhizosphère, les racines favorisent ainsi la nutrition et la santé de la plante. Améliorer les caractéristiques du système racinaire des plantes cultivées permet d’optimiser ces propriétés dans des contextes agronomiques de plus en plus contraints.
3. Le système racinaire : une cible pour l’amélioration des plantes
3.1. Diversité et complexité des systèmes racinaires
Le développement racinaire d’une même plante se développe et s’adapte de façon dynamique aux conditions environnementales. Une grande diversité dans la façon dont les systèmes racinaires se développent et s’adaptent à des environnements variés peut être observée entre individus d’une même espèce végétale. Cette diversité des systèmes racinaires entre variétés d’une même espèce est le fruit d’une sélection progressive, au fil de l’évolution, de certains caractères adaptatifs contrôlés génétiquement.
Chez le riz, des caractères d’architecture racinaire distincts ont ainsi été mis en évidence chez des variétés adaptées à des modes de culture différents :
- Des variétés adaptées à la culture de type irrigué (où le champ est en permanence recouvert d’eau) présentent un système racinaire colonisant plutôt les couches superficielles du sol ;
- Des variétés adaptées à la culture de type pluvial (où l’apport d’eau dépend seulement des pluies intermittentes) développent plutôt un système racinaire en profondeur.

Cette diversité de caractères racinaires entre plantes d’une même espèce peut être exploitée pour l’amélioration variétale. Le transfert par croisement de caractères racinaires bénéfiques d’une variété à une autre représente une stratégie d’amélioration.
Suivant ce principe, des chercheurs japonais ont pu améliorer la tolérance à la sécheresse de la variété de riz IR64. Cette variété est très productive et ses grains sont de bonne qualité. Néanmoins, elle possède un système racinaire qui se développe en surface et qui ne permet pas à la plante de prélever assez d’eau lorsque les premiers centimètres du sol s’assèchent. A l’inverse, une autre variété de riz, appelée Kinandang Patong, est moins productive mais mieux adaptée à la sécheresse grâce à la capacité de son système racinaire à puiser l’eau en profondeur. Un croisement entre ces deux variétés a permis d’introduire, chez IR64, le caractère “système racinaire profond” [18]. Une nouvelle variété de riz combinant les caractéristiques agronomiques d’IR64 au système racinaire profond et à la tolérance à la sécheresse de la variété Kinandang Patong a ainsi été produite.
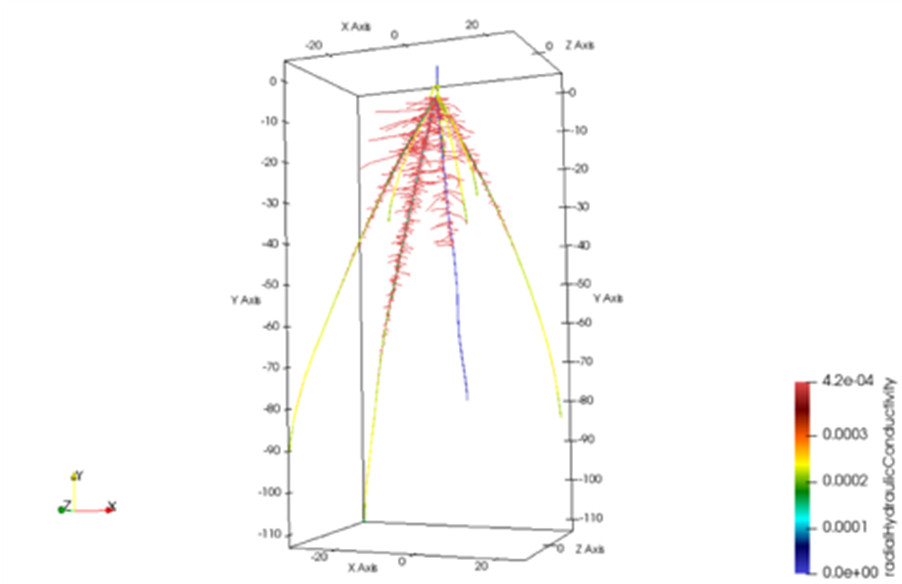
Des modèles in silico de systèmes racinaires en interaction avec des modèles de sol ont été développés et permettent de simuler la structure et les fonctions de la racine dans son ensemble (Figure 9). Ces modèles facilitent l’identification de caractères racinaires dont la modification aura, en définitive, des effets bénéfiques sur la nutrition et le développement de la plante dans un environnement donné.
3.2. Utilité des systèmes racinaires pour l’amélioration variétale
Des méthodes performantes permettent aujourd’hui de visualiser et de décrire de façon quantitative les caractéristiques du système racinaire de plantes sans les perturber. Néanmoins, ces données sont généralement collectées en laboratoire dans des conditions reflétant mal les conditions de développement des plantes cultivées au champ. Pour répondre au besoin de caractériser les systèmes racinaires de plantes en conditions agronomiques et faciliter leur prise en compte dans les programmes d’amélioration variétale, plusieurs méthodes d’étude des systèmes racinaires ont été développées.

En multipliant le nombre d’observations et de données, le développement de ces nouvelles méthodes d’exploration racinaire au champ contribue de façon décisive aux efforts menés pour :
- caractériser la diversité des caractères racinaires et leur plasticité
- et identifier les régions du génome impliquées dans le contrôle de ces caractères.
4. Messages à retenir
- Les caractères d’architecture et d’anatomie racinaires influencent la capacité des plantes à acquérir l’eau et les nutriments et à s’adapter à certains stress dont la sécheresse.
- Le développement et le fonctionnement du système racinaire répondent de façon dynamique à l’environnement. Ce phénomène appelé plasticité racinaire contribue fortement aux capacités d’adaptation de la plante à certaines contraintes.
- L’exsudation de composés carbonés par les racines dans le sol permet de modifier les caractéristiques physiques, chimiques et biologiques de la rhizosphère. Elle facilite l’acquisition de l’eau et des nutriments et intervient dans la mise en place d’interactions symbiotiques avec des microorganismes du sol.
- Toutes les plantes n’ont pas le même système racinaire. Il existe une grande diversité, entre espèces et parfois entre plantes d’une même espèce, dans la manière dont les racines se développent, fonctionnent, s’adaptent à l’environnement et interagissent avec le sol. Cette diversité, liée à des différences de patrimoine génétique entre plantes, peut être exploitée en amélioration variétale.
- Les recherches visant à améliorer les performances du système racinaire nécessitent la prise en compte de l’ensemble des caractères racinaires dans leur environnement et leur complexité. Pour cela, des méthodes de caractérisation efficaces au champ et les stratégies de modélisation sont utiles.
- La sélection variétale ciblant des caractères racinaires favorables a permis d’améliorer les performances de plusieurs espèces d’intérêt agronomique, comme le Ces recherches contribuent à une agriculture plus vertueuse, durable et économe, visant à améliorer la sécurité alimentaire des populations.
Références
Image de couverture. Racines d’arbres (1890), dernier tableau de Van Gogh (1853-1890), Musée Van Gogh, Amsterdam. [Source : Vincent van Gogh, Public domain, via Wikimedia Commons]
[1] Atkinson JA, Rasmussen A, Traini R, Voss U, Sturrock C, Mooney SJ, Wells DM & Bennett MJ. 2014. Branching out in roots: Uncovering form, function, and regulation. Plant Physiology 166, 538–550
[2] Les dessins originaux de systèmes racinaires de pois et de maïs de la figure 1 proviennent de : Kutschera, L.; Lichtenegger, E. & Sobotik, M., Würzelatlas der Kulturpflanzen gemässigter Gebiete mit Arten des Feldgemüsebaues. – Frankfurt am Main : DLG-Verlag, 2009 (2. Aufl. 2018). – 527 p. ; Wageningen University & Research Image Collections; website, Root systems drawing: https://images.wur.nl/digital/collection/coll13/search
[3] Doblas VG, Geldner N & Barberon M. 2017. The endodermis, a tightly controlled barrier for nutrients. Current Opinion in Plant Biology 39, 136–143
[4] Trinh CD, Laplaze L & Guyomarc’h S. 2018. Lateral root formation: Building a meristem de novo. Annual Plant Reviews Online 1, 847–890
[5] L’auxine fournit aux cellules une information positionnelle permettant les changements d’identité cellulaire régulant la dynamique de la morphogenèse chez les plantes. Les cellules peuvent lire la concentration de l’auxine non seulement dans l’espace mais également dans le temps. https://www.insb.cnrs.fr/fr/cnrsinfo/lauxine-permet-aux-cellules-de-lire-lespace-et-le-temps
[6] Orman-Ligeza B, Morris EC, Parizot B, et al. 2018. The Xerobranching response represses lateral root formation when roots are not in contact with water. Current Biology 28, 3165-3173.e5
[7] Motte H, Vanneste S & Beeckman T. 2019. Molecular and environmental regulation of root development. Annual Review of Plant Biology 12, 1–24
[8] de la Fuente Cantó C, Simonin M, King E, Moulin L, Bennett MJ, Castrillo G & Laplaze L. 2020. An extended root phenotype: the rhizosphere, its formation and impacts on plant fitness. Plant Journal 103, 951–964
[9] Les phytosidérophores sont des molécules de type ligand (ou chélateurs) sécrétées dans la rhizosphère par certaines espèces de plantes de la famille des Poaceae (graminées) en situation de carence en fer, et dont la fonction est de permettre l’assimilation du fer.
[10] Chen YT, Wang Y & Yeh KC. 2017. Role of root exudates in metal acquisition and tolerance. Current Opinion in Plant Biology 39, 66–72
[11] Les racines protéoïdes sont des racines de plantes qui forment des groupes denses de courtes radicelles latérales très rapprochées. Elles peuvent constituer un épais tapis de deux à cinq centimètres d’épaisseur situé juste sous la litière de feuilles mortes. Elles permettent une intense absorption des nutriments du sol, probablement en modifiant chimiquement leur environnement dans le sol pour améliorer la solubilisation des nutriments.
[12] Johnson JF, Allan DL, Vance CP & Weiblen G. 1996. Root carbon dioxide fixation by phosphorus-deficient Lupinus albus. Contribution to organic acid exudation by proteoid roots. Plant Physiology 112, 19–30
[13] Rabbi SMF, Tighe MK, Flavel RJ, Kaiser BN, Guppy CN, Zhang X & Young IM. 2018. Plant roots redesign the rhizosphere to alter the three-dimensional physical architecture and water dynamics. New Phytologist 219, 542–550
[14] Venturi V & Keel C. 2016. Signaling in the rhizosphere. Trends in Plant Science. 11, 4265–4281
[15] Phytohormones dérivées des caroténoïdes, les strigolactones contrôlent la ramification des plantes, entraînent la germination des graines de phanérogames parasites et stimulent la croissance des champignons symbiotiques mycorhiziens.
[16] Ramírez-Flores MR, Perez-Limon S, Li M, Barrales-Gamez B, Albinsky D, Paszkowski U, Olalde-Portugal V & Sawers RJH. 2020. The genetic architecture of host response reveals the importance of arbuscular mycorrhizae to maize cultivation. eLife 9, 1–18
[17] Schneider HM, Klein SP, Hanlon MT, Kaeppler S, Brown KM & Lynch JP. 2020. Genetic control of root anatomical plasticity in maize. Plant Genome 13, 1–14
[18] Uga Y, Sugimoto K & Ogawa S, et al. 2013. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nature genetics 45, 1097–102
[19] Développée par le Pr. Jonathan Lynch, cette méthode permet de noter visuellement des caractéristiques architecturales de la couronne racinaire d’une plante sur le terrain, en quelques minutes. Le terme « shovelomics » a été construit à partir du mot « shovel » (bêche en anglais) et « omics » (méthode permettant de générer des données massives et surtout de les analyser et d’en extraire les connaissances nécessaires à une meilleure compréhension du vivant).
[20] Atkinson JA, Pound MP & Bennett MJ, Wells DM. 2019. Uncovering the hidden half of plants using new advances in root phenotyping. Current Opinion in Biotechnology 55, 1–8
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Pour citer cet article : GRONDIN Alexandre, GUYOMARC'H Soazig, LAPLAZE Laurent (13 octobre 2023), Le système racinaire des plantes : de l’ombre à la lumière, Encyclopédie de l’Environnement. Consulté le 23 juillet 2026 [en ligne ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/vivant/racines-plantes/.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.


