


La quête inlassable de l’eau par les plantes
PDF
Comment l’eau parvient-elle aux feuilles des grands arbres ? Quelles astuces les plantes développent-elles face au manque d’eau ? Comment la végétation modifie-t-elle le climat ? La vie est née dans l’eau, et les plantes terrestres ont dû s’adapter à des manques d’eau temporaires. Les racines permettent de puiser l’eau du sol, les feuilles par leur transpiration exercent une succion qui tire cette eau à travers les tiges. L’eau est donc sous tension ou sous pression négative dans les tiges. Quand cette tension devient trop forte par manque d’eau, elle provoque une cavitation avec une bulle de gaz dans les vaisseaux qui cessent d’être conducteurs. Pour l’éviter, la transpiration est régulée par les stomates, qui se ferment. La fermeture stomatique limite l’entrée du CO2 dans la feuille et par suite la photosynthèse et la production.
1. Plantes et eau : les grands principes
1.1. Les plantes ont besoin d’eau liquide
Cette affirmation est triviale pour qui a chez lui quelques plantes : il faut les arroser régulièrement, certaines très souvent, d’autres moins. Tous les êtres vivants sont constitués de cellules remplies d’eau, et les plantes contiennent généralement entre 70 et 95 % d’eau. Comme toujours, il y a quelques exceptions : le bois des arbres en contient 30 à 70 %, et les graines peuvent se dessécher presque complètement, en conservant leur capacité à germer.
1.2. Les plantes ont aussi besoin d’air, et oscillent entre faim et soif
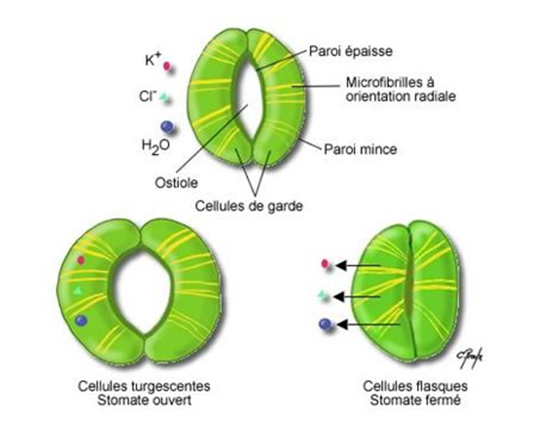
d’eau (turgescence) dans les cellules de garde, dont les parois internes sont épaissies. [Source : © Chantal Proulx]
Quand la plante manque d’eau, elle ferme ses stomates mais cela arrête aussi son alimentation en CO2, d’où un risque de mourir de faim. La plante doit donc ajuster en permanence l’ouverture de ses stomates pour réguler ses pertes en eau tout en assurant sa subsistance. Les stomates existent depuis au moins 400 Ma (millions d’années), et les plantes ont optimisé leur capacité d’échanges avec l’air extérieur depuis 325 Ma.
1.3. L’eau des plantes est-elle sous pression ou sous tension ?

débourrement, aussi appelé « pleurs » [Source : © INRAE / Montpellier SupAgro]
Cette pression de l’eau existe en permanence dans les cellules végétales et permet leur croissance :
elle pousse leurs parois, fait gonfler les cellules et augmenter leur volume et la taille des organes végétaux.
Mais lorsque les arbres ont des feuilles, il n’est pas possible de récolter de la sève, car pendant la journée l’arbre transpire et l’eau des vaisseaux est sous tension, une pression négative provoquée par l’évaporation de l’eau des feuilles. C’est la situation la plus commune pour une plante, comme nous allons le préciser.
1.4. Comment l’eau circule-t-elle dans la plante ?

Pourtant nous voyons autour de nous des arbres de 15 à 30 m de haut, qui ne semblent avoir aucun problème pour alimenter leurs feuilles en eau (Figure 3), et il existe même en Californie et en Australie des arbres atteignant 100 m de hauteur. Comment font-ils ? Cela a longtemps été discuté parce que la réponse heurte le sens commun, mais elle ne fait plus de doute aujourd’hui.
L’eau d’une feuille, présente dans les cellules et les espaces intercellulaires, s’évapore à l’intérieur de la feuille pendant la journée. Cette perte d’eau provoque une succion ou pression négative dans l’eau liquide aux sites d’évaporation. Comme les molécules d’eau sont très liées entre elles par des forces électrostatiques, cette pression négative tire l’eau du sol jusqu’aux feuilles, à travers les racines et les vaisseaux des tiges. Ainsi les plantes peuvent aspirer l’eau sur plus de 10 m de hauteur, ce que les hommes ne savent pas faire avec une pompe aspirante.
Nous avons vu que l’eau circule par gravité et sous l’effet de différences de pression. La présence de sels dissous dans l’eau attire aussi les molécules d’eau par ce qu’on appelle la force osmotique. Et lorsque l’eau se trouve dans un milieu poreux comme le sol ou la paroi cellulaire, elle se trouve liée à la matrice solide par des forces de liaison dites de capillarité. Les physiciens ont cherché à unifier ces diverses forces dans la notion de potentiel hydrique ou énergie potentielle de l’eau, l’eau circulant toujours dans le sens des potentiels décroissants. Pour préciser davantage, il nous faut détailler les propriétés physiques de l’eau, la notion de potentiel hydrique et les facteurs affectant la transpiration.
2. Plantes et eau : les détails
2.1. Les propriétés physiques de l’eau
L’eau a des propriétés remarquables liées à sa structure moléculaire, formée de deux liaisons O-H faisant un angle de 104° entre elles et constituant un dipôle électrique (Figure 4). Le côté positif de ce dipôle attire le côté négatif d’une autre molécule, assurant la cohésion du liquide. Cette force de cohésion est importante et explique la forte énergie nécessaire pour évaporer l’eau : en moyenne 590 cal/g, soit 2450 J/g ou 2,45 MJ/kg. Cette énergie, la plus élevée de tous les liquides, représente environ 6 fois l’énergie nécessaire pour porter l’eau de 0 à 100°C.
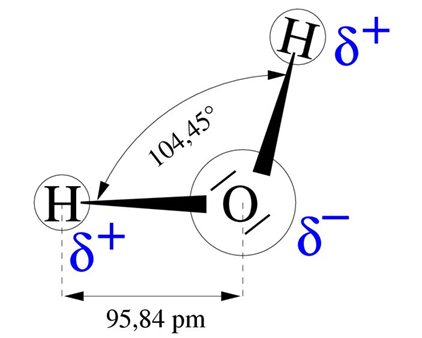
= 10-12 m. [Source : schéma de l’auteur]
Une autre conséquence de la forte cohésion entre molécules d’eau est la forte densité de l’eau liquide, supérieure à celle de la glace, ce qui permet à la glace de flotter. L’eau présente également une forte tension superficielle, qui permet aux gerridés (insectes appelés improprement araignées d’eau) de courir sur la surface des étangs.
2.2. Le potentiel hydrique et ses composantes
Le potentiel hydrique Ψ est l’énergie potentielle de l’eau, modifiée par les effets de la pesanteur, les phénomènes osmotiques et matriciels et les différences de pression (Lire Comment les plantes supportent-elles un régime salé ?). On l’exprime en joules par mètre cube (J m-3), dimensions d’une pression (en pascal, Pa). Le pascal étant une unité très petite, on utilise fréquemment le MPa ou le bar (1 bar = 0,1 MPa).
Le potentiel de gravitation Ψg est l’énergie potentielle de gravitation m.g.h, rapportée à l’unité de volume V, où m est la masse d’eau, g l’accélération de la pesanteur et h la hauteur par rapport à un niveau de référence (la surface du sol en général) :
- Ψg = (m/V).g.h = ρ.g.h
Ψg est positif si l’eau est au-dessus du sol (h>0), et négatif sinon (h<0). Ainsi dans un arbre, Ψg est négatif pour l’eau des racines et positif pour celle des feuilles.
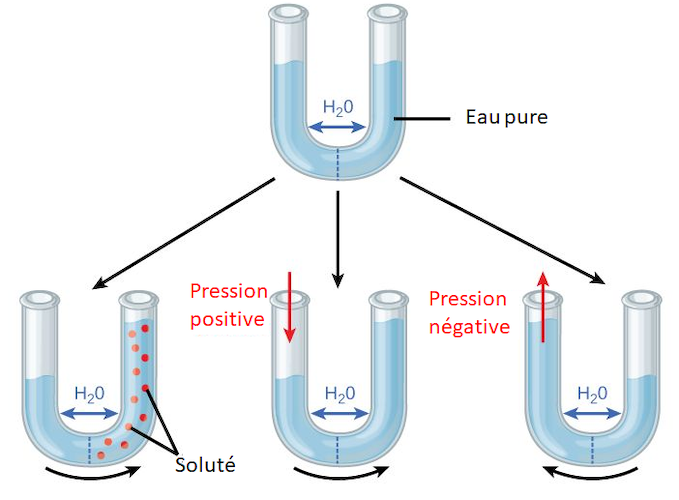
Le potentiel osmotique Ψs est toujours négatif car l’eau liée aux solutés est moins disponible que l’eau libre. Il dépend de la concentration de solutés Cs. En première approximation pour une solution diluée, cette dépendance est linéaire :
- Ψs = – R.T. Cs
où R est la constante des gaz parfaits et T la température absolue
Le potentiel matriciel (Ψm) est l’énergie liant l’eau aux particules solides de la matrice (sol ou paroi cellulaire), il est proportionnel à la surface de contact (4πR2 pour un pore sphérique), et rapporté au volume (4 πR3/3), il varie comme 1/R, soit comme l’inverse du rayon du pore. Il est toujours négatif, et d’autant plus que les pores sont petits. Ainsi un sol sableux à pores grossiers retient peu l’eau, tandis qu’un sol argileux à pores très petits la retient bien davantage. L’eau du sol disponible pour les plantes est comprise entre deux limites :
- Une limite supérieure : la capacité au champ ;
- et une limite inférieure : le point de flétrissement permanent.
Au-delà de la capacité au champ, l’eau s’écoule par gravité, en deçà du point de flétrissement permanent, l’humidité résiduelle est non accessible aux plantes et correspond à Ψm = – 15 bars (ou – 1.5 MPa).
On appelle réserve utile l’intervalle entre ces deux limites, pour un sol de 1 m de profondeur, elle représente entre 100 et 200 mm de hauteur d’eau, ce qui représente 30 à 60 jours de consommation pour une culture en bonnes conditions. En pratique, les plantes commencent à fermer leurs stomates quand environ la moitié de cette réserve a été utilisée, et la réserve est d’autant plus faible que la profondeur de l’enracinement est inférieure à 1 m. Mais les arbres ont des racines bien en dessous de 1 m, augmentant donc la réserve utile. Elle atteint 400 mm dans la forêt de chêne sessile de Barbeau près de Melun, d’après les chercheurs qui l’étudient au laboratoire d’Écophysiologie végétale d’Orsay (voir ci-dessous).
Enfin, le potentiel de pression Ψp est directement égal à la pression, et donc positif dans une cellule turgescente et négatif dans les vaisseaux pendant la journée.
Le potentiel hydrique Ψ est donc la somme de ces quatre composantes, et l’eau circule toujours dans le sens des potentiels décroissants :
- Ψ = Ψg + Ψs + Ψm + Ψp
La Figure 6 illustre les variations de ce potentiel dans un arbre transpirant pendant la journée :
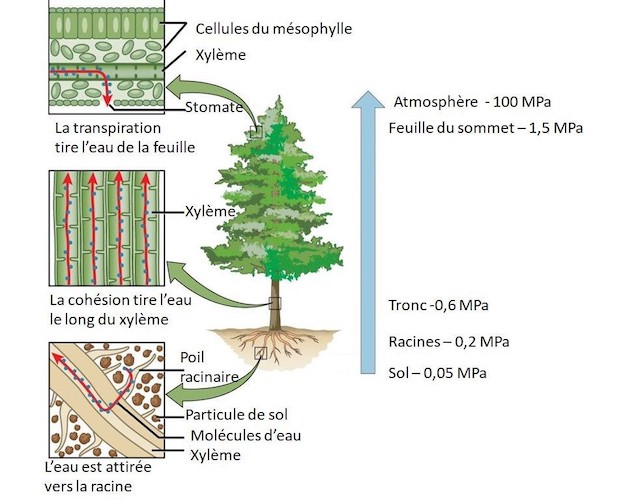
- Le potentiel du sol est légèrement négatif (- 0,05 MPa soit – 0,5 bar) parce que l’eau y est en dessous de la surface et qu’elle contient des sels minéraux dissous en faible concentration.
- En haut de l’arbre, le potentiel d’une feuille est de – 1,5 MPa ou -15 bars, car la feuille est soumise à une transpiration importante. Cette valeur fortement négative exerce une succion sur toute la colonne d’eau, des racines au feuillage, ce qui donne des valeurs intermédiaires pour les racines (-0,2 MPa) et pour le tronc (-0,6 MPa). La colonne d’eau supporte ces valeurs négatives de potentiel hydrique (et donc de pression) grâce aux forces de cohésion qui relient entre elles les molécules d’eau.
- On notera que le potentiel de l’air (-100 MPa) est très inférieur à celui de la feuille (-1,5 MPa). Ce potentiel est en équilibre avec l’humidité relative de l’air, ici 48% à l’air libre, contre 98,9% dans l’air interne à la feuille, ce qui est donc presque saturé. [2]
La hauteur maximale des arbres est de l’ordre de 100 m, ce qui correspond à un potentiel gravitationnel de 10 bars ou 1 MPa. Il faut donc que les feuilles soient à un potentiel nettement inférieur à – 1 MPa pour pouvoir faire monter l’eau jusqu’à cette hauteur.
2.3. Évaporation et couche limite, résistance aérodynamique
L’eau est présente sous forme de vapeur dans l’air. Lorsqu’on injecte de la vapeur d’eau dans de l’air sec, la pression de vapeur d’eau (e) augmente jusqu’à une valeur maximale, au-delà de laquelle la vapeur se condense en eau liquide. C’est ce qu’on appelle la pression de vapeur saturante es(T), qui augmente de façon exponentielle avec la température (Figure 7). Le rapport e/ es(T) est l’humidité relative et s’exprime en %. La différence es(T) – e est le déficit de saturation, qui augmente fortement avec la température.
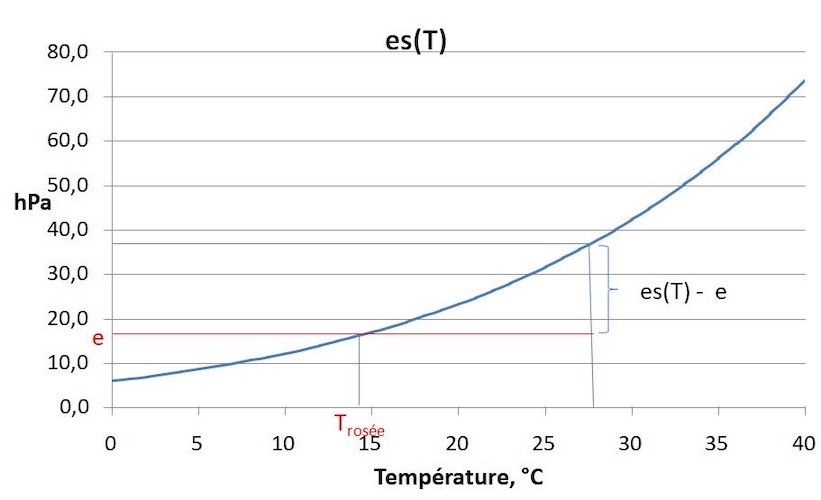
L’air au contact avec de l’eau liquide est saturé en vapeur d’eau, et l’évaporation d’une surface d’eau libre E va
dépendre du déficit de saturation [es(T) – e] entre la surface et l’air libre. E augmente avec la vitesse du vent, et on définit une résistance aérodynamique ra, résistance opposée par l’air au flux de vapeur d’eau E entre la surface évaporante et l’air libre.
L’essentiel de cette résistance est situé dans ce qu’on appelle la couche limite, qui va de la surface où la vitesse de l’air est nulle, à l’air libre. Son épaisseur est de l’ordre du millimètre pour une feuille, de quelques centimètres pour une surface d’eau libre, et peut atteindre plusieurs décimètres pour un champ de maïs et plusieurs mètres pour une forêt.
2.4. Transpiration
La résistance ra peut s’appliquer à une surface d’eau libre (mare, lac) ou à un objet mouillé comme un papier buvard humide. Pour une plante, la vapeur d’eau doit d’abord diffuser à travers l’épiderme et ses stomates, avant de sortir dans l’air extérieur, ce qu’on représente par rs, une résistance qui s’ajoute à ra comme
deux résistances en série.

On compare souvent une feuille à un papier buvard humide qui évapore librement sous l’effet des facteurs physiques. Le rayonnement solaire apporte l’essentiel de l’énergie d’évaporation, la vitesse du vent modifie la résistance ra, température et humidité de l’air affectent le déficit de saturation de l’air. L’évaporation est d’autant plus grande que le rayonnement est élevé, et l’air chaud et sec. La différence entre la feuille et le buvard humide, c’est la présence de l’épiderme et de ses stomates, qui ralentit l’évaporation, comme une feuille de plastique percée de petits trous :
- Quand les stomates sont bien ouverts, la transpiration est proche de l’évaporation du buvard (un peu moins tout de même) ;
- Quand les stomates se ferment, la transpiration diminue jusqu’à des valeurs très faibles liées à la porosité résiduelle de la cuticule de l’épiderme. Il en résulte une augmentation de la température foliaire qui n’est plus refroidie par l’évaporation.

et calculée à l’aide de modèles physiques.
2.5. Évapotranspiration
Le flux E au-dessus d’un couvert végétal s’appelle évapotranspiration, c’est la somme de l’évaporation du sol et des surfaces mouillées (s’il y en a), et de la transpiration des feuilles. Il dépend étroitement de l’indice foliaire, qui est le rapport entre la surface de feuilles d’une végétation et la surface de sol sous-jacente.
En pratique, le flux d’eau évaporée par une surface naturelle dépend des conditions de la surface (eau libre, sol plus ou moins couvert de végétation et plus ou moins humide) et des conditions climatiques (rayonnement solaire, température et humidité de l’air, vitesse du vent).
On appelle évapotranspiration potentielle Ep la quantité d’eau évaporée par une surface de végétation basse couvrant entièrement le sol et bien alimentée en eau. Ep varie de 0 à 10 mm/j selon les climats ; sur une année Ep peut aller jusqu’à 2700 mm, valeur maximale observée dans des endroits chauds et secs comme au Sahel, pour de l’herbe irriguée bien verte.

Nationale]

ces mesures depuis 2005 au-dessus d’une forêt de chênes près de Fontainebleau (Figure 11) [4].
3. Rôle de l’eau dans la production végétale
3.1. La transpiration est-elle nécessaire aux plantes ?
Oui, car les feuilles ont besoin d’être ouvertes sur l’air extérieur pour pouvoir absorber le dioxyde de carbone dont elles se nourrissent, et c’est la raison pour laquelle leur épiderme est percé de nombreux stomates (Lire Quel est l’impact des polluants de l’air sur la végétation ?). L’air interne à la feuille est très humide car il est en contact avec des cellules bien hydratées, et on ne connaît pas de cellules végétales ou animales pouvant fonctionner sans eau. La transpiration résulte de cette porosité de l’épiderme foliaire et de la différence d’humidité entre l’air intérieur et l’air extérieur à la feuille. Les plantes ont aussi besoin d’éléments minéraux autres que le carbone (azote, phosphore, potassium…) qu’elles trouvent dans la solution du sol absorbée par les racines grâce à la transpiration.
On considère que pour produire un kilo de blé, une culture doit consommer environ 500 L d’eau. Ce chiffre est un ordre de grandeur, il est un peu plus faible en climat humide et plus élevé en climat aride à cause des variations d’évapotranspiration.
3.2. Conséquences pour la circulation de la sève
L’eau liquide est formée de molécules polaires, qui s’attirent les unes les autres avec des forces de cohésion particulièrement élevées. Cette grande cohésion permet la montée de la sève dans les grands arbres dont l’eau des vaisseaux subit des pressions négatives fort basses. Toutefois, la colonne d’eau des vaisseaux peut se rompre lorsque la tension devient trop forte, une bulle de gaz apparaît alors et bloque la circulation d’eau, c’est la cavitation, qui est l’équivalent de l’embolie pour les vaisseaux sanguins. Un arbre comporte de nombreux vaisseaux, et commence à souffrir quand la cavitation frappe une part importante de ces vaisseaux [6]. L’anatomie des vaisseaux en relation avec leur sensibilité a été beaucoup étudiée.
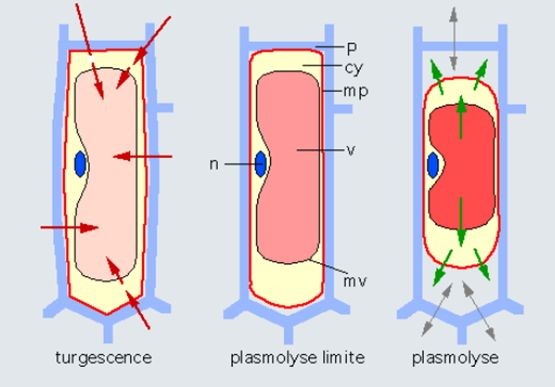
On trouve l’eau d’une plante dans les cellules qui la composent, et aussi dans les vaisseaux qui alimentent la plante des racines jusqu’aux feuilles. L’eau du sol est en général peu chargée en éléments minéraux, alors que l’eau des cellules est assez concentrée. Cette différence de concentration attire par osmose un flux d’eau du sol vers la plante, qui va gonfler les cellules (Figure 13, à gauche). L’eau est donc normalement sous pression dans les cellules, on dit que les cellules sont turgescentes. Cette pression est contenue par les parois cellulaires, constituées de fibres de cellulose et de pectine relativement rigides, qui agissent comme un pneu qui contient la pression d’une chambre à air. Un pneu de voiture est gonflé entre 2 et 3 bars, un pneu de vélo peut aller jusqu’à 6 bars, et la pression peut aller jusqu’à 10 bars dans une cellule végétale. C’est en raison de ces pressions élevées que les cellules végétales ont une paroi, qui n’existe pas dans les cellules animales, soumises à des pressions
plus faibles. Cette pression diminue en cas de dessèchement, puis s’annule, la membrane de la cellule se décollant alors de la paroi, c’est la plasmolyse (Figure 13, au centre et à droite).
4. Messages à retenir
- L’eau est le milieu nécessaire à toutes les formes de vie.
- Elle est sous pression dans les cellules végétales, et cette pression permet la croissance et assure la forme et la rigidité des plantes, avec aussi les tissus ligno-cellulosiques des tiges et des troncs.
- Par contre, elle est sous tension dans les feuilles et les vaisseaux, et cette tension permet la montée de la sève, qui compense les pertes d’eau des feuilles par transpiration et évaporation.
- La transpiration a lieu principalement à travers les stomates, ces pores nécessaires pour assurer l’absorption du dioxyde de carbone.
- Les stomates sont bien ouverts sur sol humide et se ferment sur sol sec pour conserver l’eau interne, freinant alors l’entrée du CO2 et donc la photosynthèse et la croissance.
- La fermeture stomatique est déclenchée par la baisse du potentiel hydrique qui, lorsqu’elle devient forte, provoque aussi la formation de bulles de gaz dans les vaisseaux par cavitation.
Les mécanismes d’adaptation des plantes au manque d’eau sont traités dans l’article associé (à paraître).
Notes et références
Image de couverture. Culture de vigne dans le parc de La Geria sur l’île de Lanzarote (Canaries). Les rares pluies (170 mm/an) sont retenues par les murets de pierre et donnent un vin de qualité. [Source : Patrice78500, CC BY-SA 4.0, via Wikimedia Commons]
[1] https://fr.wikipedia.org/wiki/Eau_de_bouleau
[2] La formule liant le potentiel à l’humidité atmosphérique est : Ψ = (RT/Veau)*ln(e/es(T))
où R est la constante des gaz parfaits, T la température absolue, Veau le volume d’une mole d’eau liquide et e/es(T) l’humidité relative. En prenant R = 8,3 J mol-1 K-1, T = 298 K, Veau = 18.10-6 m3 mol-1 et e/es(T) = 0,48 on obtient Ψ = – 100 MPa. Ainsi le potentiel hydrique de l’air est en général très bas, sauf quand l’air est presque saturé (hr>98%) comme dans l’air interne à la feuille
[3] Dufrêne E, Pontailler J-Y et Saugier B, 1993. A branch bag technique for simultaneous CO2 enrichment and assimilation measurements on beech (Fagus sylvatica L.) Plant, Cell and Environment, 16, 1131-1138
[4] http://www.barbeau.universite-paris-saclay.fr/index-fr.html
[5] Jung M et al. (2010). Recent decline in the global land evapotranspiration trend due to limited moisture supply. Nature 467:951-954.
[6] Le lecteur intéressé trouvera plus d’informations dans l’excellente présentation d’André Granier et Hervé Cochard dans le site « Les plantes et l’eau : II.7. L’architecture hydraulique, la cavitation et l’embolie des plantes ».
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Pour citer cet article : SAUGIER Bernard (11 août 2022), La quête inlassable de l’eau par les plantes, Encyclopédie de l’Environnement. Consulté le 18 juin 2026 [en ligne ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/vivant/quete-de-eau-par-les-plantes/.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.


