


Inheritance or convergence? The winding paths of species evolution
PDF
How can we explain that some species are more alike than others? Species are the product of their history: evolution. They represent the leaves of the tree of life, which emerges from a main trunk symbolizing their ancestor common to all. Two closely related species in this tree have therefore recently separated from each other – they have a recent common ancestor – and often resemble each other. Two species far away in the tree are therefore generally very different because they have followed independent evolutionary branches. However, some pairs of historically distant but very morphologically similar species are counter-examples. In fact, these intriguing similarities are the product of the repeated adaptation of species to the same environmental conditions. These evolutionary convergences are major elements that make it possible to affirm that adaptation is a fundamental force in the evolution of living organisms.
A remarkable feature of the living world is the very great diversity of forms it takes. Individuals are often grouped into species, which makes it easy to describe this biodiversity. Thus, a certain diversity of species forms within mammals can be observed in the images in Figure 1. But some species are more alike than others. Thus, the polar bear looks much more like the brown bear than the gorilla.
1. First cousins are more alike than distant cousins
Species have long been described from a fixistAssumption that there is neither transformation nor drift of living species, nor is there any profound modification of the Universe. point of view, i.e. as immutable entities, “essences” that were a priori thought by the divine (see What is biodiversity?). At the end of the 18th and beginning of the 19th centuries, scientists such as Lamarck (1744-1829), Wallace (1823-1913) and Darwin (1809-1882, see focus on Darwin) revolutionized this traditional vision of biodiversity by proving that living species evolved from their origins (see Lamarck and Darwin: two divergent visions of the living world) and that they were therefore all descended from a single common ancestor. How does this discovery help us to better understand the similarities between species?
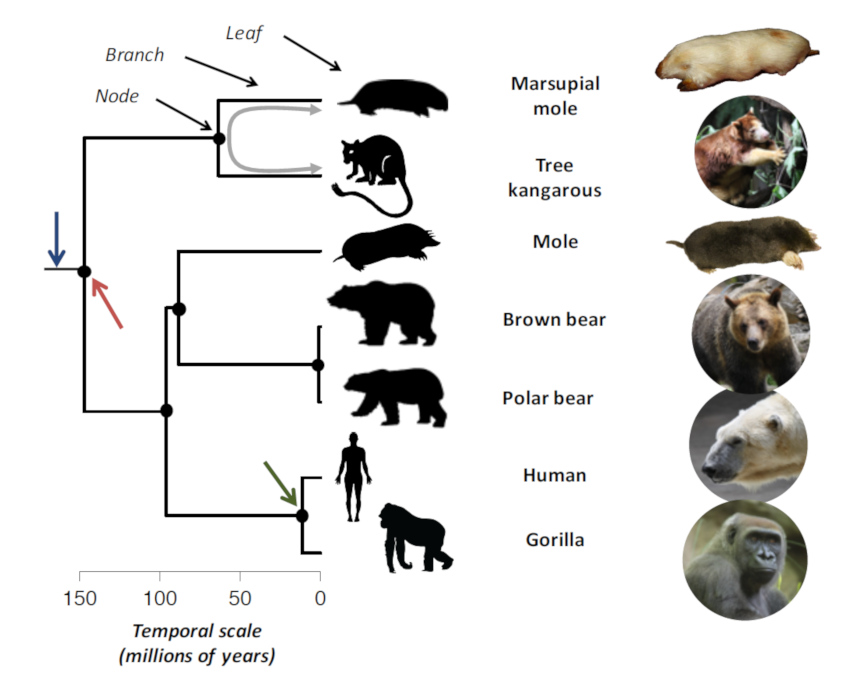
2. Some counter-examples…
However, there are counter-examples: evolutionarily distant species can look strikingly similar, as is the case between some large groups of marine animals, plants or mammals.
2.1. Marine animals
Radiated finfish (e.g. tuna) [4] and cetaceans (marine mammals including whales and dolphins) separated about 430 million years ago [5]. However, don’t they share a similar general appearance? On this subject, Darwin [6] already noted that:
” For animals, belonging to two most distinct lines of descent, may readily become adapted to similar conditions, and thus assume a close external resemblance; but such resemblances will not reveal -will rather tend to conceal their blood-relationship to their proper lines of descent. […] thus the shape of the body and fin-like limbs […] being adaptations in both classes for swimming through the water…”
Darwin suggests here that both groups have, over evolution, developed a hydrodynamic form and fins that allow them to move efficiently in the water.
2.2. Underground mammals
Mole-like animals (Figure 1) exist in mammals in different parts of the world. Thus, the European mole is present in part of Eurasia while the marsupial mole is present in Australia. These species have a similar burrowing shape, adapted to underground life (Figure 1). They all have a stocky “sausage-shaped” body – without a distinguishable neck – equipped with powerful forelegs with large claws for efficient digging. They have all more or less lost their external sensory organs (external ear, sight). The similarity between these “moles” does not come from a common heritage, that is, from a common ancestor with a mole shape. Indeed, the marsupial mole is phylogenetically closer to a kangaroo than to a European mole (Figure 1), which is closer to man than the marsupial mole. Thus, the “mole” form has appeared several times in evolution of mammals.
2.3. Cushion plants

2.4. “Cactoid” plants

2.5. Carnivorous plants

3. When natural selection “blurs the tracks”
Thus, although some species may be very distant phylogenetically, they may have converged towards a similar morphology, to adapt to similar environmental constraints. The hypothesis of a random evolution that took place previously (hypothesis 1) therefore does not seem to be able to explain these phenomena of adaptation of living organisms to their environment. A major contribution of Darwin (1859) was precisely to propose the mechanism allowing the adaptation of species: natural selection.
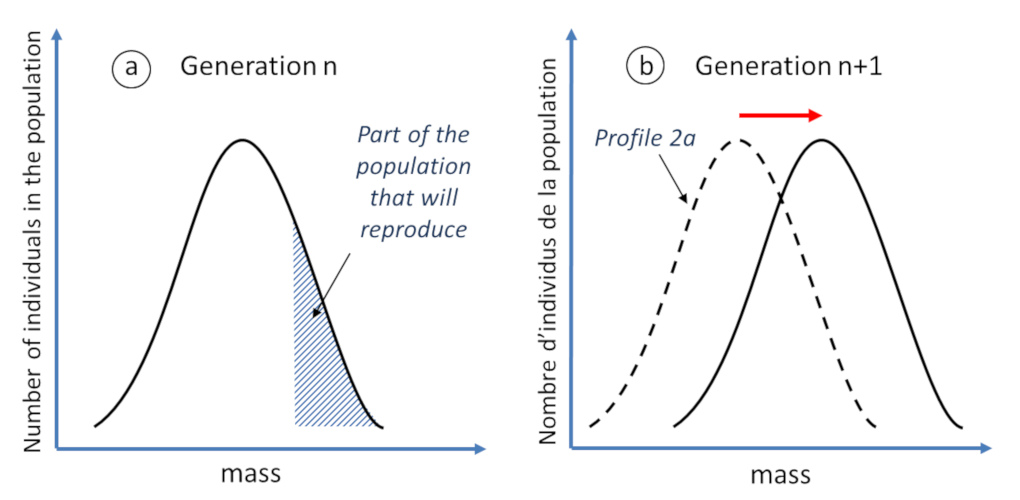
In Darwin’s theory, natural selection requires four ingredients:
- The first necessary condition is the existence of a certain diversity of forms within a species itself, for example individuals of more or less compact cushion plants (Figure 5A). It is precisely this diversity that was ignored by the fixists, for whom it represented only “errors of copying” the divine essence of the species. However, this diversity is real: just look at the diversity within the human species.
- The second condition is limited reproduction of individuals. Just imagine that each individual of a species gives two descendants to each generation. If we start from a single individual, after twenty generations, the population will have reached the size of 220 individuals, i.e. more than one million and after a hundred generations, several billion of billions… which is a priori impossible on a planet with important but limited resources. Thus, only a limited number of individuals of the species will contribute to the next generation.
- The third condition stipulates that the set of individuals belonging to the breeding lot has particular characteristics that have allowed them to better survive in an environment with given constraints and therefore to reproduce more. For example, in an increasingly harsh environment, it is conceivable that only the most compact cushion plant individuals survive long enough to reproduce (Figure 5A).
- If these characteristics are heritable, then they will gradually spread throughout the population and the species will gradually evolve over generations (Figure 5B).
Darwin provides here a theory to explain the adaptation of species to their environment. Thus, if two independent branches of the tree are subject to the same environmental constraints – such as nitrogen deficiency or harsh climate – they can lead to morphologically similar but phylogenetically distant species. In short, species are mosaics of characters that may have taken different evolutionary paths. For example, all cushion plants have the same general shape, but have very distinct floral morphologies, characteristic of their family (Figure 3). Cetaceans have lungs – inherited from a distant vertebrate ancestor – packed in a hydrodynamic morphology that is not inherited, but convergent.
4. Synthesis
Similarities between species can therefore be the result of historical heritage or evolutionary convergence. To be able to distinguish between the two hypotheses, it is therefore necessary to have a phylogeny of the species and to measure their morphological, anatomical or physiological characteristics. Historically, phylogenetic trees were built from morphological or anatomical data, which could be problematic if converging characters were used to build them. In this case, there was a risk of grouping together species that have in fact converged and whose similarities are not inherited. The last few decades have seen the advent of DNA molecule sequencing and its use in phylogeny. Molecular similarities are much less subject to these evolutionary convergences, and if properly used they are therefore a tool of choice for reconstructing phylogenetic relationships between species. The comparison between the old taxonomic classificationsRelative to taxonomy, or hierarchical classifications of living beings. based on morphology and phylogenetic classifications from molecular data will highlight evolutionary convergences. It is, in a way, a synergy between old and new approaches in evolution.
References and notes
Cover image. Dolphin group photo made in Xcaret, Mexico. [Source: © Truncatus [public domain] via Wikimedia Commons]
[1] Darlu P. & Tassy P. (1993) Phylogenetic reconstruction. Concepts and Methods. Series “Theoretical Biology”, No. 7. Masson, Paris (online book on the website of the Société Française de Systématique: http://sfs.snv.jussieu.fr/index.php/la-reconstruction-phylogenetique-concepts-et-methodes/).
[2] Bininda-Emonds O.R., Cardillo M., Jones K.E., Jones K.E., MacPhee R.D., Beck R.M., Grenyer R., Price S.A., Vos R.A., Gittleman J.L. & Purvis A. (2007) The delayed rise of present-day mammals. Nature, 446, 507-512.
[3] The dates mentioned in this article are from the study by Bininda-Edmonds et al (reference 2). These dates are speculative and reflect the current state of knowledge in mammalian phylogeny, and are therefore subject to modification as new discoveries are made.
[4] “Fish” includes cartilaginous fish (sharks and rays), radiated finfish (e.g. trout or tuna), hagfish, lampreys and lungfish. They no longer form a group used in systematics (but – rightly – still in cooking), because the common ancestor of “fish” is not exclusive to them. For example, the ancestor common to trout and shark is also the common ancestor of man. The “fish” do not therefore represent an evolutionary lineage of their own, but rather a concatenation of several distinct groups.
[5] http://www.timetree.org. These dates are speculative and reflect the current state of knowledge in phylogeny, and are therefore subject to modification as new discoveries are made.
[6] Darwin C. (1859) On the origins of species by means of natural selection. London: Murray, 247.
[7] see also www.cushionplants.eu
The Encyclopedia of the Environment by the Association des Encyclopédies de l'Environnement et de l'Énergie (www.a3e.fr), contractually linked to the University of Grenoble Alpes and Grenoble INP, and sponsored by the French Academy of Sciences.
To cite this article: MAZEL Florent, LAVERGNE Sébastien, DOUZET Rolland, THUILLER Wilfried (January 5, 2025), Inheritance or convergence? The winding paths of species evolution, Encyclopedia of the Environment, Accessed July 29, 2026 [online ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/en/life/inheritance-or-convergence-winding-paths-of-species-evolution-2/.
The articles in the Encyclopedia of the Environment are made available under the terms of the Creative Commons BY-NC-SA license, which authorizes reproduction subject to: citing the source, not making commercial use of them, sharing identical initial conditions, reproducing at each reuse or distribution the mention of this Creative Commons BY-NC-SA license.


