




继承或是趋同?物种进化的蜿蜒之路

我们如何解释一些物种彼此之间比其他物种更相似的问题?物种来源于其历史:进化。它们代表着生命之树的叶子,而生命之树则是从代表着它们共同祖先的主干上生长出来的。因此,这棵树上的两个密切相关的物种,他们拥有最相近的祖先并且往往彼此相似,直到最近才将彼此分离。因此,在树上相距甚远的两个物种一般非常不同,因为它们遵循着独立的进化分支。然而,却有一些进化历史相异,但在形态上非常相似的物种成为了例外。事实上,这些耐人寻味的相似之处是物种反复适应相同环境条件的结果。这些进化上的趋同使得我们能够肯定适应是生物进化基本力量的主要因素。
生物界的一个显著特点是其形式极其多样。个体常被归为物种,这使得描述这种生物多样性变得容易。因此,在图1中可以观察到哺乳动物内部物种形式的某种多样性。但有些物种之间比起其他物种更为相似。比起大猩猩,北极熊和棕熊看起来更加相像。
1. 表亲比远亲更相似
长期以来,人们一直以固定的视角描述物种 ,即物种是不可改变的实体,是由神灵先验地决定的“本质”(见“什么是生物多样性?”)。在18世纪末、19世纪初,拉马克(Lamarck,1744-1829)、华莱士(Wallace,1823-1913)和达尔文(Darwin,1809-1882,见焦点“达尔文”)等科学家彻底改变了这种看待生物多样性的传统观点。他们证明了生物物种是由其祖先演变而来的(见“拉马克和达尔文:生命世界的两个不同视角”),因此,它们都是同一个共同祖先的后代。这一发现是如何帮助我们更好地理解物种之间相似性的呢?
Branch:分支
Node:节点
Temporal Scale (millions of years):时间尺度 (百万年)
Marsupial mole: 有袋类鼹鼠
Tree kangarous:树袋熊
Mole:鼹鼠
Brown bear:棕熊
Polar bear:北极熊
Human:人类
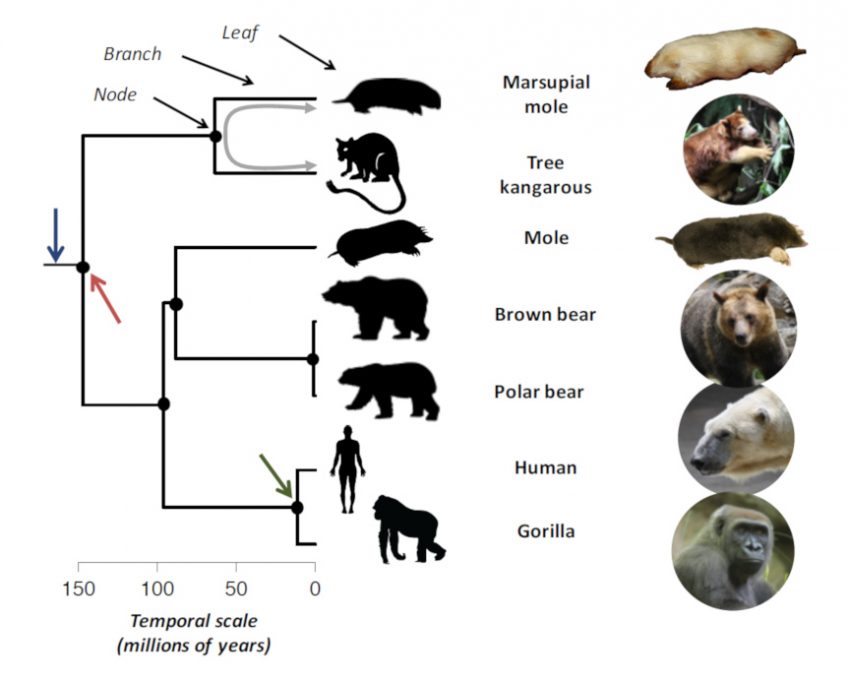
Gorilla:大猩猩 图1. 系统发育树的一个实例。(现有近5000个哺乳动物物种中的)七个物种为系统发育树的叶子,而系统发育树描述了它们的进化关系。水平比例尺以百万年为单位。蓝色箭头表示树的根,红色箭头表示它的第一个节点,这也是树上所有物种的共同祖先(1.47亿年前)。绿色箭头表示大猩猩和人类的共同祖先,它们被认为生活在1150万年前。双灰色箭头代表树袋鼠和有袋类鼹鼠之间的系统发育距离(共同祖先年龄的两倍,即2×63 = 1.26亿年)。该系统发育树引自比宁达·艾德蒙兹(Bininda-Edmonds)等人的研究[2]。
[来源:© F·梅泽尔 欧洲鼹鼠除外〔迪迪埃·德库昂(Didier Descouens)〕和有袋类鼹鼠〔迈克尔·J·巴里特(Michael J. Barritt)〕]
物种的进化可以表示为一棵系统发育树 (生物群体间亲缘关系的示意图。树的每个节点代表其后代的共同祖先。它所代表的名称是由兄弟群体组成的支系的名称,而不是祖先的名称,因为后者仍然无法确定。树可能有根,也可能没有,这取决于是否已经确定了所有叶子节点共同的祖先。)(图1),它重建了“有组织生命的后裔的历史进程”[1]。他凭此描述了物种之间的相互关系。树上的每个分支都有叶子,即现存的物种。树的每个节点都代表它以下物种的共同祖先。这棵树标了时间刻度——通常以百万年为单位——时间从根部开始流动(从图1中的蓝色箭头开始由左向右)[2]。根代表祖先物种,是树上所有叶子的共同祖先。在图1的例子中,它在约1.47亿年前分为两个子进化枝(红色箭头)[3],这两个子进化枝在到达现存的物种——叶子之前,本身也有几次分支。大猩猩和人代表了两片近缘的叶子,也就是说,两者之间的共同祖先(绿色箭头),比人和,例如,北极熊之间的共同祖先相比,历史更近。如果物种以恒定的速率随时间随机进化(假设1),那么它们的相似度就会因此降低,因为它们在很久以前就分开了。例如,人和大猩猩的差异只积累了1150万年,而人和北极熊的差异却积累了9100万年,这反映了和北极熊相比,大猩猩和人之间的相似性非常高。因此,物种之间的系统发育距离 (表示两个分类单元之间的整体相似性指数。这是序列及其祖先之间进化的程度。灰色箭头)可以很好地说明两个物种之间预期的形态相似性:两个物种在系统发育上的关系越密切,它们就越有可能彼此相似。
2. 一些反例……
然而,也有相反的例子:进化上相距甚远的物种也可以看起来惊人地相似,如一些海洋动物、植物或哺乳动物之间的情况。
2.1. 海洋动物
辐鳍鱼(如金枪鱼)[4]和鲸类(海洋哺乳动物,包括鲸鱼和海豚)在大约4.3亿年前分离[5]。然而,它们看上去不是很像吗?对于这个问题,达尔文[6]已经指出:
“属于两个差异最大谱系的动物,可能很容易因为适应类似的条件而呈现出密切的外部相似性;但这种相似性不会揭示——反而会掩盖它们与真实谱系的血缘关系。……因此,身体的形状和鳍状肢体……在两类动物中都是用于适应在水中游泳的。”
达尔文在这里提出,这两个群体在进化过程中出现了流线型身体和鳍状肢,这使得它们能够在水中高效地移动。
2.2. 地下哺乳动物
世界不同地区的哺乳动物中均存在类鼹鼠动物(图1)。欧洲鼹鼠生活在欧亚大陆的部分地区,而袋鼹鼠则生活在澳大利亚。这些物种都具有类似利于掘洞的身形,适应于地下生活(图1)。它们都有一个粗壮的 “香肠形”身体——没有明显的颈部——并配备了强大的前肢,有大爪子,可以有效地挖掘。它们都或多或少地失去了外部感觉器官(外耳、视觉)。这些“鼹鼠”之间的相似性并非来自具有鼹鼠外形的共同祖先的继承。事实上,袋鼹鼠在系统发育上比欧洲鼹鼠更接近袋鼠(图1),而欧洲鼹鼠比有袋动物鼹鼠更接近人类。因此,“鼹鼠”的形态在哺乳动物的进化过程中独立出现过几次。
2.3. 垫状植物

[来源:照片 © 约瑟夫·富里埃高山植物园(Joseph Fourier Alpine Station,SAJF)] 。
垫状植物是在一定程度上形成紧凑圆顶状的植物(图2[7])。这种极其特殊的形态是由非常狭窄的节间(两个连续的叶子之间的茎)和小的叶片造成的。在法国,这些生物体可以在气候恶劣、寒冷多风的高山上找到。事实上,这种植物的结构非常紧凑,不仅可以保护根和茎不受温度骤变的影响,而且还能抵御寒冷和大风,使植物能够在极端条件下生存。可以看出在历史上植物曾多次进化为垫状,因为垫状植物多次出现在差异极大的科中,并出现在世界所有具有极地气候或高山气候的地区(图2)。
2.4. “类仙人掌”植物

[来源:照片 © 塞巴斯蒂安·桑特(Sébastien Sant)和 ©让-保罗·佩尔蒂埃(Jean-Paul Pelletier), www.teline.fr]
类仙人掌植物,即外形类似仙人掌的植物,是生活在干旱地区的具有特征性外观的植物(图3)。它们的叶子有时会变成刺,使它们得以限制水分的流失,并保护自己免受食草动物的侵害。此外,它们都含有储存水分的组织。在美洲的沙漠中,拥有这种特殊形态的是仙人掌属的植物,而在非洲,大戟属和犀角类植物[1]取代了它们(图3)。这三类植物属于独立的进化群体,因此这种极其特殊的形态并非继承而来,而是在进化过程中独立获得的。
2.5. 食虫植物

[来源:照片。分别由卡斯滕·尼豪斯(Carsten Niehaus)、诺亚·艾尔哈特(Noah Elhardt)和安德烈斯·艾尔斯(Andreas Eils)拍摄,自维基共享资源网]。
食虫植物是能够吸引和捕捉昆虫等小动物的植物(图4)。在法国,它们通常被发现于缺氮的环境中,如泥炭沼泽 (被植被占据的湿地,其特殊的生态条件促使土壤形成,土壤中含有非常高的有机物质,几乎没有或很少植物来源的分解。泥炭地是一种特殊而脆弱的生态系统,尽管存在甲烷排放,泥炭地的特点使其成为一个碳汇,因为有机物的合成比降解更多。见“泥炭地与沼泽,非凡的湿地”)。食虫植物通过消化这些被捕获的动物大量补充氮元素,从而弥补了土壤中可用氮的缺乏。在系统发育关系相距较远的科中分别发现了这种植物,表明在植物的进化过程中,食虫的特征多次独立出现。虽然,这些物种从动物身上获得氮元素的功能很明显是趋同进化的,但为实现这一功能而发育的结构却更加多变。例如:昆虫可以被粘性物质固定(图4A和4B),这种粘性物质也在进化中反复出现;或者被困在一个瓮状结构中(图4C)。在后一种情况下,植物发育了不同的结构来捕捉动物,这正是著名的“进化修补”的例证:多种结构可以用于实现同样的功能。
3. 当自然选择“模糊了来路”
因此,尽管一些物种的系统发育关系可能非常遥远,但它们可能已经趋同于类似的形态,以适应类似的环境限制。之前提到的随机进化的假设(假设1)似乎不能解释生物体适应环境的现象。达尔文(1859)的一个主要贡献正是
提出了使物种得以适应环境的机制:自然选择。
在达尔文的理论中,自然选择有四个必要条件:
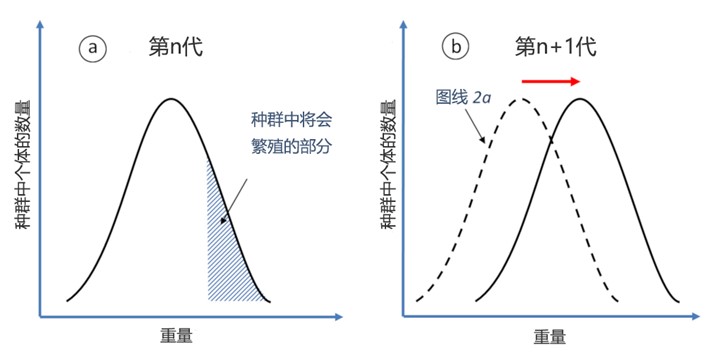
- 第一个必要条件是一个物种本身存在着某种形式的多样性,例如垫状植物个体或高或低的紧凑程度(图5a)。固定论者恰恰忽视了这种多样性,对他们来说,这种多样性只代表物种神圣本质的“复制错误”。然而,这种多样性是真实存在的:只要看看人类的多样性就能明白这一点。
- 第二个条件是个体繁殖受限。想象一下,假如一个物种的每个个体在每一代都会有两个后代。如果我们从一个个体开始,经过20代,人口将达到220个个体的规模,即超过100万;而经过100代,将达到几十亿的规模……这在一个资源重要而有限的星球上根本是不可能的。因此,该物种只有有限的个体能对下一代做出贡献。
- 第三个条件规定,同一批被繁殖出的个体具有独特的特征,使它们能够在有某种限制的环境中更好地生存,因此能够繁殖更多的后代。例如,可以想到,在一个日益恶劣的环境中,只有最紧凑的垫状植物个体才能存活足够长的时间,并得以繁殖(图5a)。
- 如果这些特征是可遗传的,那么它们将逐渐在整个种群中扩散,物种将一代代逐渐进化(图5b)。
达尔文在这里提出了一个理论来解释物种对环境的适应。因此,如果发育树上的两个独立分支受到了相同的环境限制——如氮缺乏或恶劣气候——就会出现形态上相似但系统发育上相距甚远的物种。简而言之,物种像是从不同可能的路径进化来的特征组成的马赛克贴画。例如,所有的垫状植物形状大致相同,但其花的形态各异,仍具有各自的科的特征(图3)。鲸类具有肺,这是从遥远的脊椎动物祖先那里继承来的,但却包裹在趋同进化而来(不是遗传而来)的流线型身体中。
4. 总结
因此,物种之间的相似性可能是遗传继承或趋同进化的结果。为了区分这两种情况,所以有必要对物种进行系统发育分析,并测量其形态学、解剖学或生理学特征。在历史上,系统发育树是根据形态学或解剖学数据建立的,如果用趋同的特征建立系统发育树,可能会出现问题。在这种情况下,有可能将实际上是趋同进化而来的物种归为一类,而这些物种的相似性并不是通过继承得到的。过去几十年来,DNA分子测序技术得到发展,并在系统发育研究中得到应用。分子数据的相似性受进化趋同的影响要小得多,因此如果使用得当,它们是重建物种之间系统发育关系的首选工具。基于形态学的旧分类学 (taxonomic classifications)和基于分子数据的系统发育分类学之间的比较将使得趋同进化现象更加明显。在某种程度上,它也是进化中的新旧方法之间的协作。
参考资料及说明
封面照片:摄于墨西哥坎昆的海豚群。[来源:© Truncatus [公共领域]自维基共享资源网]
[1] Darlu P. & Tassy P. (1993) Phylogenetic reconstruction. Concepts and Methods. Series “Theoretical Biology”, No. 7. Masson, Paris (online book on the website of the Société Française de Systématique: http://sfs.snv.jussieu.fr/index.php/la-reconstruction-phylogenetique-concepts-et-methodes/).
[2] Bininda-Emonds O.R., Cardillo M., Jones K.E., Jones K.E., MacPhee R.D., Beck R.M., Grenyer R., Price S.A., Vos R.A., Gittleman J.L. & Purvis A. (2007) The delayed rise of present-day mammals. Nature, 446, 507-512.
[3] 本文中提到的时间点来自比宁达·艾德蒙兹(Bininda-Edmonds)等人的研究(参考文献2)。这些时间点仅为推测,反映了我们目前对哺乳动物系统发育关系的认知,因此,随着新发现出现,这些时间点会被修正。
[4] “鱼”包括软骨鱼(鲨鱼和鳐鱼)、辐鳍鱼(如鳟鱼或金枪鱼)、盲鳗、七鳃鳗和肺鱼。它们不构成系统学中定义的一个类群(但在烹饪中仍然正确地使用),因为“鱼”并不存在一个独有的共同祖先。比如,鳟鱼和鲨鱼的共同祖先也是人类的共同祖先。因此,“鱼”并不代表它们自己的进化谱系,而是几个不同群体的合并。
[5] http://www.timetree.org。这些时间点仅为推测,反映了我们目前对系统发育关系的认知,因此,随着新发现出现,这些时间点会被修正。
[6] Darwin C. (1859) On the origins of species by means of natural selection. London: Murray, 247.
[7] 另见www.cushionplants.eu
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: MAZEL Florent, LAVERGNE Sébastien, DOUZET Rolland, THUILLER Wilfried (2024年3月14日), 继承或是趋同?物种进化的蜿蜒之路, 环境百科全书,咨询于 2026年8月3日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/inheritance-or-convergence-winding-paths-of-species-evolution/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。

