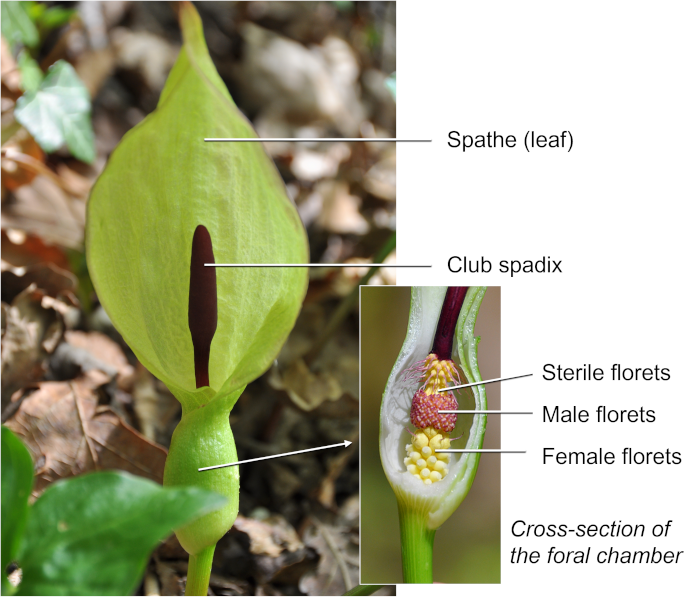
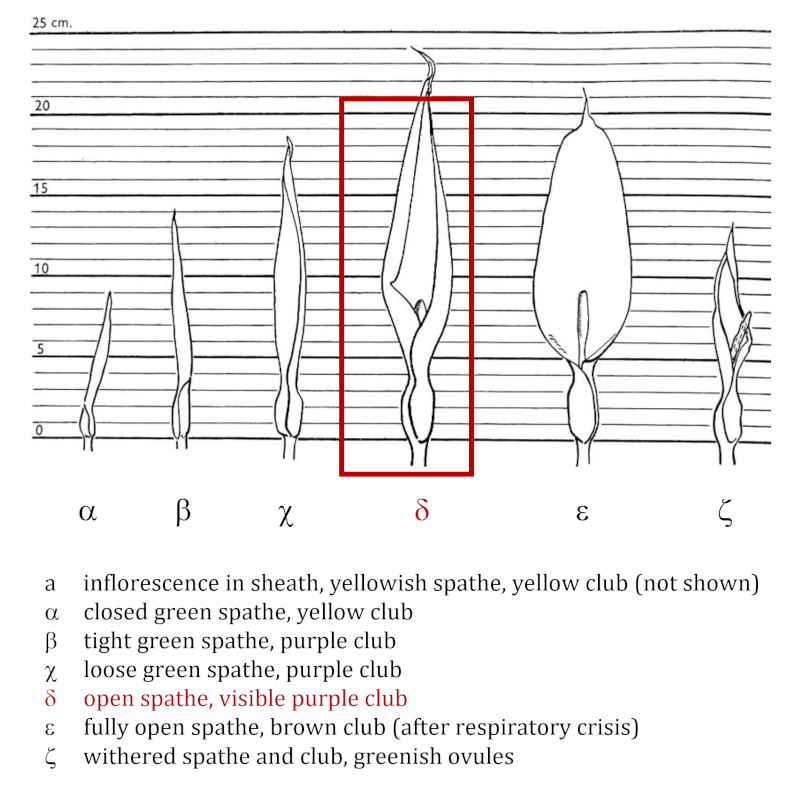
Figure 2. Representation of the various stages of development of Arum maculatum. It is during stage d (inset) that the respiratory crisis and associated heat production occurs. (Author’s diagram adapted from James and Beevers [2] and Lance [3]).The respiratory crisis of the spadix and, more specifically, the club, commonly occurs shortly after the spathe opens, as early as the δ stage. It uses starch stored during development and lasts for a few hours during which the respiratory intensity and temperature of the spadix follow a parallel evolution [4] (Figure 3). The temperature of the spadix can thus reach 30°C and be nearly 20°C above air temperature.
2. Cyanide insensitive respiration coupled with heat production
The cyanide resistance of Araceae respiration, which is particularly important at the time of respiratory crisis, was observed as early as the 1950s [2]. Isolated spadix mitochondria exhibit electron transport using both the classical cyanide-sensitive cytochromic pathway terminated by the cytochrome oxidase complex and a cyanide-resistant, non-phosphorylating, non-cytochromic pathway, the latter of which is particularly active during the respiratory crisis [5].
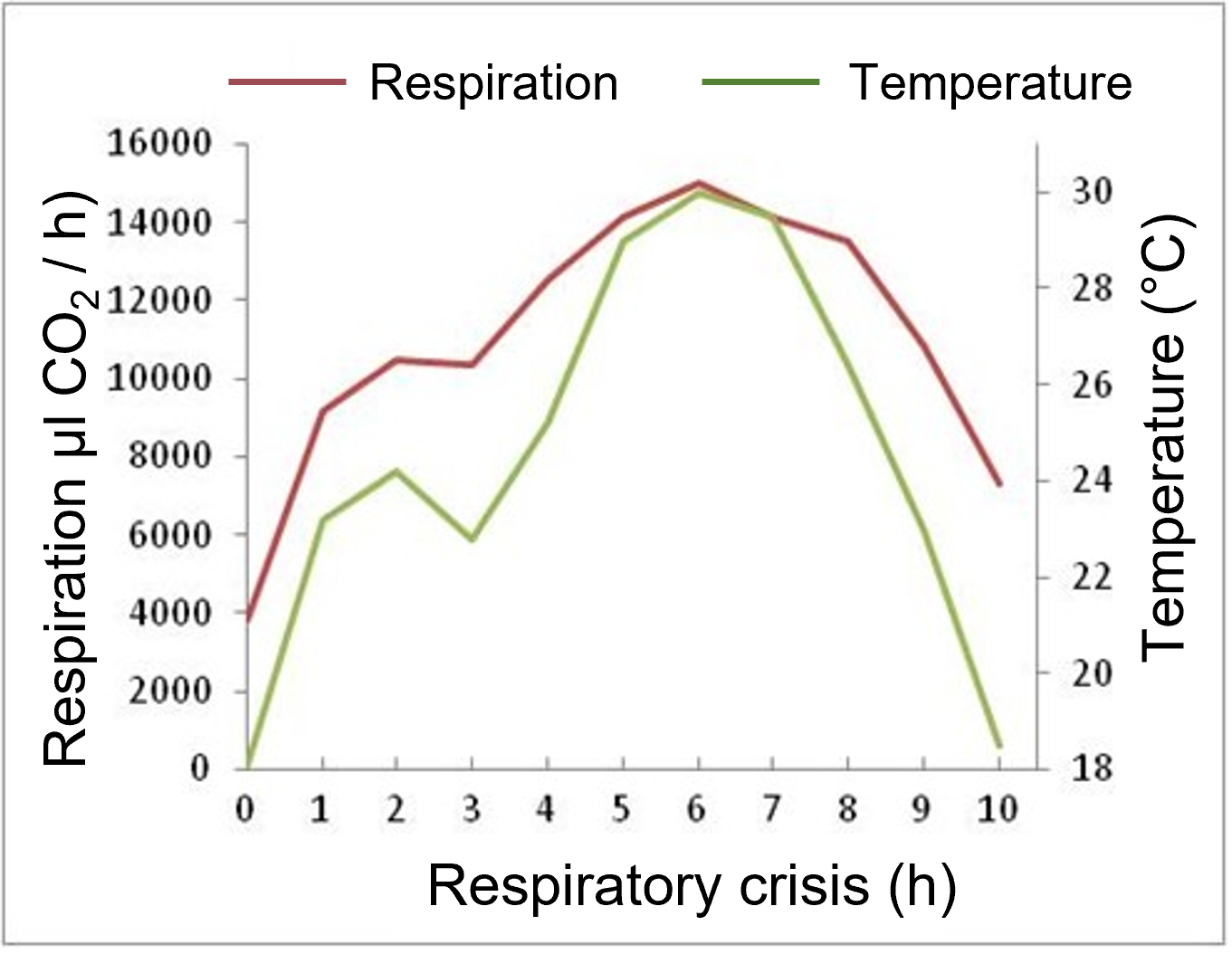
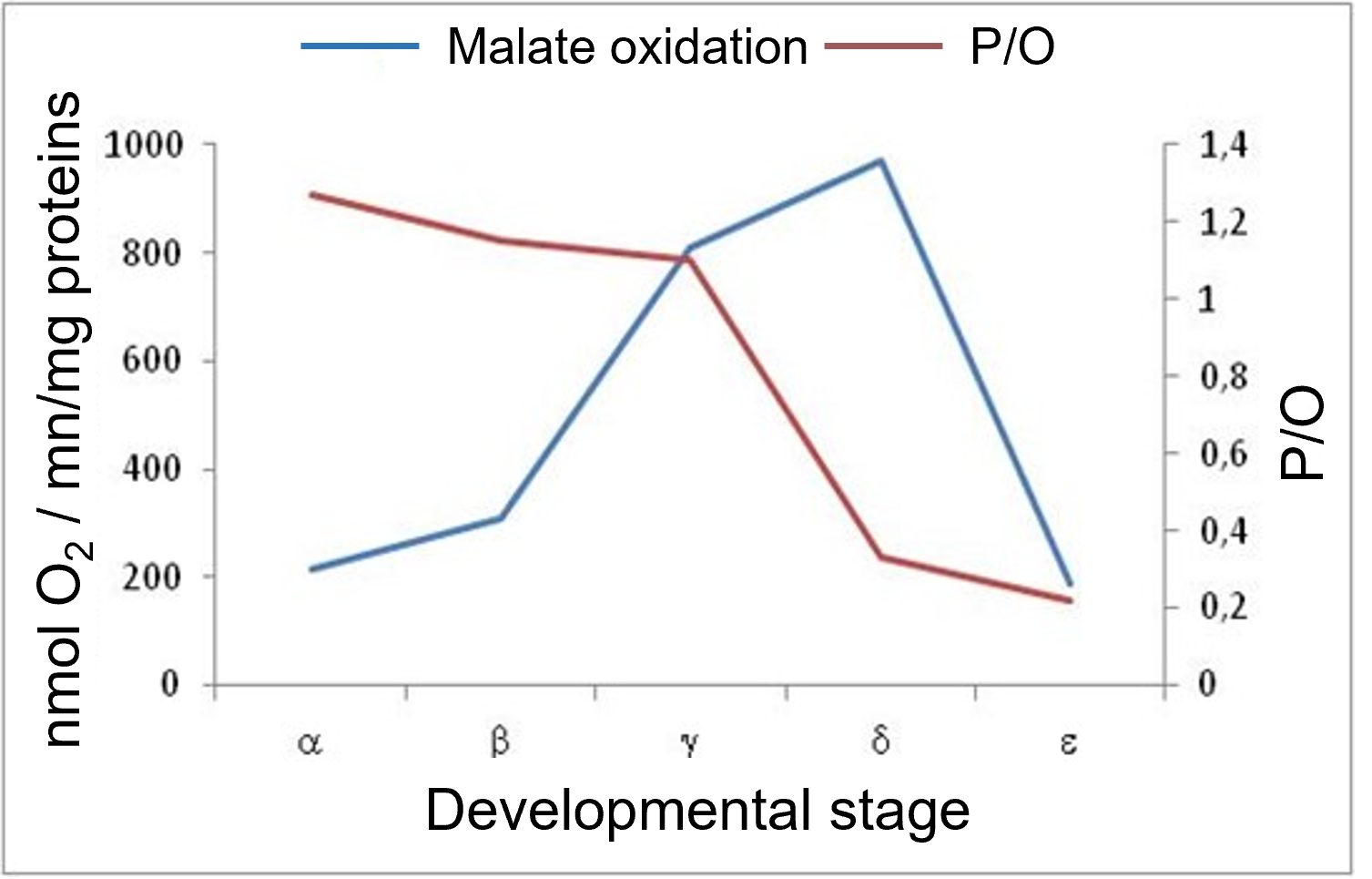
Figure 3. Changes in respiratory intensity and spadix temperature at the time of respiratory crisis [Source: Author’s diagram, adapted from ap Rees et al. [4]).Figure 3 shows the evolution of mitochondrial oxidation of malate (organic acid at the origin of NADH reoxidized by the respiratory chain, see Figure 10 of the main article) and associated phosphorylation during the developmental stages of Arummaculatum spadix [6]. Malate oxidation peaks at the time of the respiratory crisis, which corresponds to the increase in respiration. At the same time, the efficiency of oxidative phosphorylation (P/O), which corresponds to ATP production, already low in the early stages of development, decreases sharply during the respiratory crisis, indicating a significant involvement of the non-phosphorylating electron transport pathway [6]. The involvement of AOX, the terminal oxidase of this pathway, was subsequently demonstrated [7].
The low level of oxidative phosphorylation reflects the release of excess energy from the high level of respiration in the form of heat.
3. Heat production and pollination in Arum
Figure 4. Evolution, over developmental stages, of malate oxidation and oxidative phosphorylation in mitochondria of Arum maculatum clubs (Author’s diagram, adapted from Chauveau and Lance [6]).Female Arummaculatum flowers become functional (i.e., susceptible to pollination) when the inflorescence begins to open. Within the spadix, thermogenesis has started: it promotes (i) the activity of enzymes catalyzing the synthesis of volatile amino compounds and (ii) the emission of these molecules in the vicinity of Arum plants. Their odour of excrement and urine attracts pollinating insects.
Pollination of the Arummaculatum flower is carried out by small 2-3 mm midges known as psychodes (Psychoda phalaenoides) [8] and also nicknamed “sink midges” or “butterfly flies” (Figure 5). These insects are very common in damp places (e.g., sewage pipes).
Figure 5. The “most read in the burrows” newspaper, The Hoot, described in detail the process that takes place in the heart of the inflorescence of the wild arum and that attracts a species of insect, Psychoda phalaenoides (or Psykoda), thus involuntarily participating in the pollination of the Arum. [Source: Reproduced from issue 26 (1985) of “la Hulotte” (DR)]Moreover, the high temperature (15 to 20°C above room temperature) found within the spathe is very suitable for this small insect. Psychoda lands on the cone, slides, because the walls are very smooth, and passes without hindrance through the sterile flowers. These flowers are so made that they trap Psychoda in the floral chamber: filaments pointing downwards (see Figure 1) prevent the insect from going up and coming out. It is then trapped in the lower part of the inflorescence (Figure 5) [9]. Many other Psychoda insects will join it and will also remain trapped: the Arum inflorescence is a real insect trap (Figure 6) [10] !
But things end up working out for Psychoda: after 2 days, the male flowers mature, open and let out pollen which sprinkles the midges, the sterile flowers wither and open the floral chamber… The imprisoned midges can then escape… They fly to other Arums which they will then be able to fertilize, in return for a new episode of imprisonment!
Acknowledgements.We are very grateful to Mr. Laurent Francini (Association La Chanterelle de Ville-la-Grand, Haute-Savoie) for the pictures which allowed to illustrate this focus on the pollination of Arum maculatum by the midge Psychoda phalaenoides.
[1] L. Garreau, 1851. De la respiration chez les plantes, Mémoire sur les relations qui existent entre l’oxygène, consommé par le spadice de l’Arum italicum en état de paroxysme, et la chaleur qui se produit, Ann. Sci. Nat. Ser. III, 15, 5-36. (in French)
[2] W.O. James & H. Beevers, 1950. The respiration of Arum spadix. A rapid respiration, resistant to cyanide, New Phytol. 49, 353-374. https://doi.org/10.1111/ j.1469-8137.1950.tb05160.x.
[3] Lance C 1972. Respiration of Arum maculatum during inflorescence development. Annals of Natural Sciences. Botany12th series, Volume XIII, 477-495.
[4] ap Rees T, Wright BW & Fuller WA (1977). Measurements of starch breakdown as estimates of glycolysis by the spadix of Arum maculatum L. Planta, 134, 53- 56. https://doi.org/10.1007/BF00390094.
[5] James WO, & Elliott DC (1955). Cyanide-resistant mitochondria from the spadix of an arum. Nature, 175, 89. 10.1038/175089a0
[6] Chauveau M & Lance C (1982). Respiration and thermogenesis in Araceae. Bulletin de la Société Botanique de France. Actualités Botaniques. 129, 123-134. https://doi.org/10.1080/01811789.1982.10826566 (in French)
[7] Grant NM, Miller RE, Watling JR & Robinson SA (2008). Synchronicity of thermogenic activity, alternative pathway respiratory flux, AOX protein content, and carbohydrates in receptacle tissues of sacred lotus during floral development. Journal of Experimental Botany, 59, 705-714.
[11] Wagner AM, Krab K, Wagner MJ & Moore AL (2008). Regulation of thermogenesis in flowering Araceae: the role of the alternative oxidase. Biochimica Biophysica Acta, 1777, 993-1000. https://doi.org/10.1016/j.bbabio.2008.04.001