


Lumière, vision et horloges biologiques
PDF
Depuis que la vie est apparue sur Terre, diverses forces de sélection ont influencé l’évolution des organismes vivants. Parmi ces forces, la lumière générée par notre soleil a probablement eu l’influence la plus importante. En particulier, la lumière et ses cycles naturels -journaliers et saisonniers- ont conduit les organismes vivants à se représenter leur environnement dans l’espace et le temps. Ils ont ainsi pu s’adapter à cet environnement. Comment ? Quels systèmes photorécepteurs visuels et non-visuels ont émergé au sein du règne animal ? Comment les avantages fournis par ces systèmes, la vision et les horloges biologiques, ont-ils permis aux organismes de contrôler leur environnement et d’anticiper ses changements ?
Notre étoile, le Soleil, est âgée de 4,6 milliards d’années. C’est un véritable réacteur nucléaire : chaque seconde, en son cœur, 620 millions de tonnes d’hydrogène sont converties en 616 millions de tonnes d’hélium. Les quatre millions de tonnes manquantes sont converties en énergie rayonnée vers l’extérieur, c’est-à-dire en photons, autrement dit en lumière. Chaque seconde, après un trajet de 8 minutes à la vitesse de 300 000 km par seconde, 6 400 mille milliards de photons par millimètre carré pénètrent l’atmosphère terrestre. Une partie d’entre eux est absorbée ou réfléchie vers l’espace par les atomes et les molécules qui la composent. L’autre partie atteint la surface terrestre et illumine notre planète.
Depuis que la vie est apparue sur Terre, il y a près de 3,8 milliards d’années, 1 400 milliards de cycles jour/nuit et 3,8 milliards de cycles saisonniers se sont succédé (lire Focus Cycles de la lumière). La lumière a ainsi fortement influencé l’évolution du monde vivant avec l’apparition de systèmes photosensibles, visuels ou non-visuels, et d’horloges biologiques. En d’autres termes, la lumière et ses cycles ont permis aux organismes vivants de se représenter l’espace et le temps et donc de s’y adapter [1].
1. Histoire évolutive de la photoréception
1.1. Les protéines photoréceptrices
Différentes étapes ont marqué l’évolution des systèmes sensibles à la lumière, de l’apparition des premières protéines photosensibles aux systèmes visuels complexes actuels. Une classe importante de protéines photosensibles est à l’origine de la photoréception : la classe des opsines. Il en existe deux super-familles, les opsines de type I et II [2]. Les opsines de type I, également appelées opsines microbiennes, se retrouvent dans les trois domaines du Vivant (Archae, Bacteria et Eukarya). Elles régulent principalement les réactions d’orientation des organismes libres par rapport à la lumière, ou phototaxie, ainsi que la production d’énergie. Quand sont apparues ces opsines de type I ? Leur distribution générale dans tous les organismes suggère que leur précurseur existait déjà avant la séparation des trois domaines du Vivant. Cette origine lointaine démontre l’importance fondamentale de la lumière pour le Vivant. Les opsines de type II, également appelées opsines animales, se retrouvent uniquement chez les organismes eucaryotes supérieurs. Elles régulent principalement la photoréception visuelle ainsi que les horloges biologiques. Le précurseur de ce type d’opsine serait apparu il y a 700 millions d’années chez les métazoaires primitifs, les premiers organismes multicellulaires mobiles et hétérotrophes. La structure de ces deux types d’opsines présente des similarités, pourtant il n’existe aucun lien phylogénétique entre ces deux super-familles. Au contraire, elles auraient évoluées indépendamment l’une de l’autre [2].
1.2. Les cellules photoréceptrices
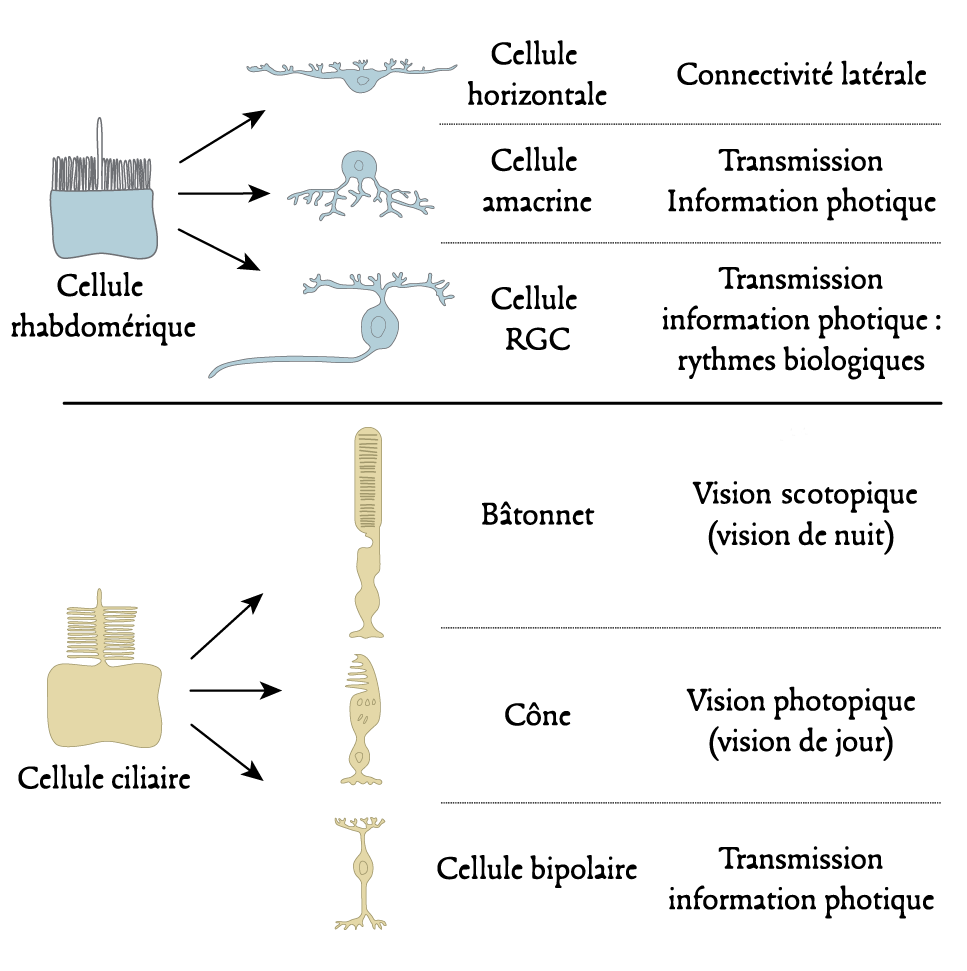
1.3. Les systèmes non-visuels et visuels
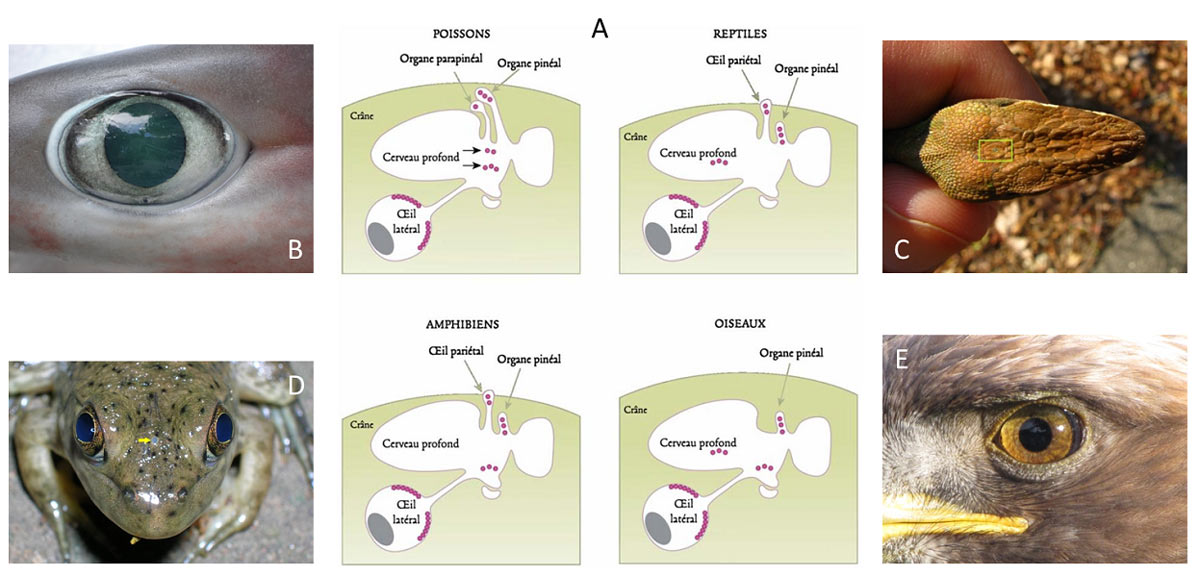
De nos jours, la plupart des animaux possèdent des systèmes non-visuels et visuels complexes. Parmi les systèmes non-visuels, on distingue diverses structures :
– un organe situé à l’intérieur du crâne, en forme de petite pomme : l’organe pinéal ; il est photosensible chez tous les vertébrés sauf les mammifères ;
– un organe associé à l’organe pinéal, d’où son nom d’organe parapinéal, intracrânien chez les poissons ;
– un organe frontal ou œil pariétal chez les amphibiens et les reptiles ;
– des photorécepteurs du cerveau profond chez tous les vertébrés, sauf les mammifères ;
– des photorécepteurs dermiques, présents sur la peau chez plusieurs espèces de vertébrés et d’invertébrés, en particulier chez les amphibiens et les céphalopodes.
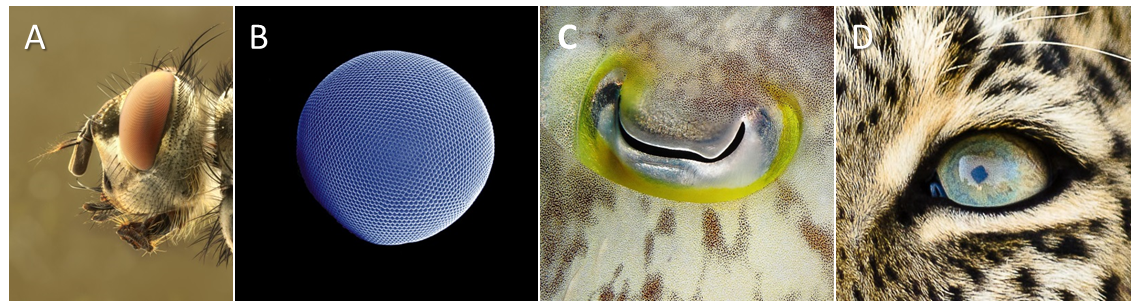
Les systèmes visuels désignent les yeux latéraux et se retrouvent chez la plupart des invertébrés et chez tous les vertébrés. On en distingue dix catégories structurellement différentes. Parmi les plus remarquables, on trouve les yeux composés des insectes et les yeux à lentille simple des vertébrés et des mollusques (Figure 3).
2. La vision
Chez tous les vertébrés, les yeux latéraux possèdent une structure commune. Elle est comparable à celle d’un appareil photo à lentille simple : la lumière traversant la pupille (la lentille) forme une image inversée sur la rétine (la surface sensible) qui tapisse la face postérieure de l’œil. La rétine est un feuillet de tissu composé de cinq types de neurones connectés entre eux : photorécepteurs visuels, cellules horizontales, cellules bipolaires, cellules amacrines (dépourvues d’axones) et cellules ganglionnaires (Figure 4). On y distingue communément une partie interne et une partie externe [5] (Figure 4).
Il existe deux formes de photorécepteurs visuels, les cônes et les bâtonnets. Les premiers sont sensibles aux fortes intensités lumineuses et sont donc impliqués dans la vision de jour, dite photopique. Les seconds ne réagissent qu’aux faibles intensités lumineuses et sont donc impliqués dans la vision de nuit, dite scotopique. La diversité de ces photorécepteurs est très grande, aussi bien au sein d’une même espèce qu’entre espèces différentes. Or, chaque type de cônes et de bâtonnets contient une protéine photoréceptrice bien précise qui n’est sensible qu’à certaines longueurs d’onde du spectre électromagnétique. Chaque espèce animale possède donc une vision monochromatique ou polychromatique qui lui est spécifique (voir plus bas). Ainsi, les poissons, amphibiens, reptiles et oiseaux sont, pour la plupart, tétrachromates : ils traitent l’information visuelle à l’aide de quatre types de photorécepteurs. Les mammifères sont en général dichromates, très probablement à cause de leur histoire évolutive associée aux niches écologiques nocturnes. Un certain nombre de primates, tel l’être humain, constituent une exception notable, car ils sont trichromates [3]. Cette propriété, nouvelle au sein des mammifères, est apparue suite à la duplication d’un gène (lire Focus Évolution de la vision).
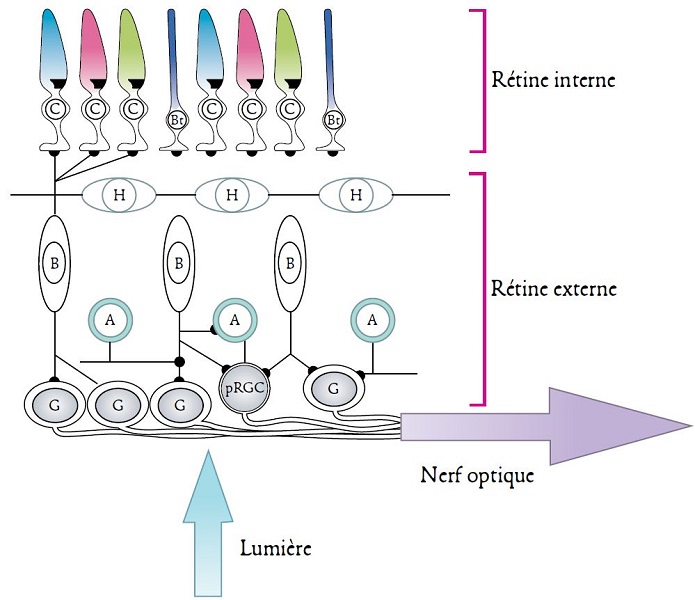
Ainsi, les organismes sont capables de percevoir les informations visuelles qui émanent de leur environnement et peuvent donc s’en faire une représentation spatiale.
3. Horloges et rythmes biologiques
3.1. Les rythmes biologiques et le cycle jour/nuit
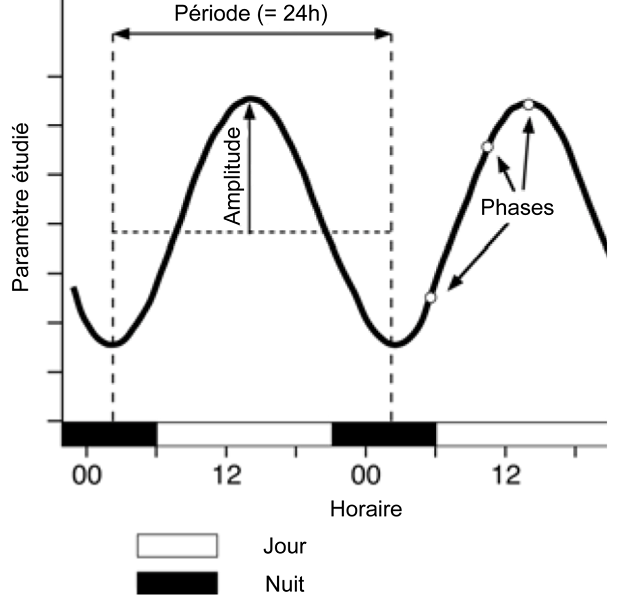
Le terme « rythme biologique » désigne un phénomène qui, au sein d’un organisme vivant, se répète selon une période et une amplitude constantes au fil du temps. Pour une espèce donnée, ces rythmes sont déterminés génétiquement et générés par une horloge interne qui est active même en l’absence de stimuli extérieurs. Il est possible de distinguer trois types de rythmes biologiques :
– les rythmes circadiens, également appelés rythmes journaliers, qui ont une période proche de 24h ;
– les rythmes ultradiens, qui ont une période inférieure à 24h ;
– les rythmes infradiens, qui ont une période supérieure à 24h ; le rythme circannuel, proche d’une année terrestre, entre dans cette dernière catégorie [6].
3.2. Horloge centrale et horloges périphériques
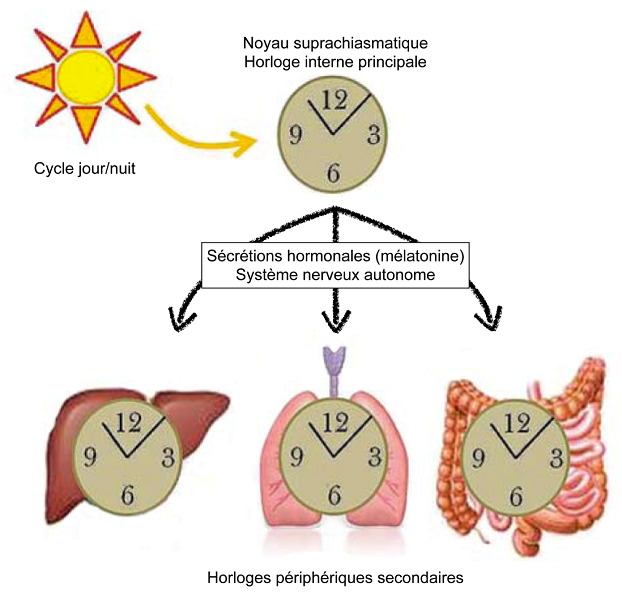
Chez les mammifères, l’horloge interne principale réside dans le noyau suprachiasmatique de l’hypothalamus. Ce noyau est composé d’environ 20 000 neurones capables d’activer spontanément et de manière cyclique des gènes « horloges ». Sans stimuli extérieurs, et en fonction des espèces, l’expression de ces gènes oscille sur une période endogène comprise entre 23h et 25h. Une fois activés, ces gènes horloges contrôlent à leur tour l’expression d’autres gènes. Au total, chez les mammifères, près de 20% du génome sont sous le contrôle de l’horloge interne principale [7].
Il existe aussi des horloges périphériques secondaires. En effet, des gènes horloges s’expriment dans la plupart des tissus périphériques. Cependant, sans l’horloge interne principale, les cycles d’expression des gènes horloges diffèrent d’un tissu périphérique à un autre. Le noyau suprachiasmatique est donc nécessaire pour synchroniser ces horloges secondaires.
Grâce à l’alternance jour/nuit, l’horloge interne principale est strictement synchronisée sur une période de 24h. Chez les mammifères, les informations photiques sont perçues par la rétine interne des yeux latéraux. Comme nous l’avons déjà vu, celle-ci est composée de plusieurs familles de cellules photoréceptrices, dont les cellules ganglionnaires photosensibles aux longueurs d’onde comprises entre 460 et 480 nm (bleues) impliquées dans la transmission d’informations non-visuelles. Ce sont ces cellules qui transmettent, via la voie rétino-hypothalamique, les informations photiques au noyau suprachiasmatique. Ces informations activent ou inhibent l’expression des gènes horloges des neurones qui composent le noyau. Dès lors, celui-ci est synchronisé par le cycle jour/nuit [8].
4. La lumière naturelle et ses rôles auprès du Vivant
4.1. La lumière naturelle et les écosystèmes
Au cours d’une journée de 24h, l’intensité lumineuse perçue par les organismes vivants varie de plusieurs magnitudes. Autrement dit, à chaque heure de la journée correspond une intensité lumineuse spécifique : le cycle jour/nuit fractionne ainsi l’environnement. Voilà pourquoi, chez les espèces animales, il existe différents profils d’activités journalières : activités diurne, nocturne, crépusculaire et cathémérale (aussi bien le jour que la nuit). Ce fractionnement a donc de fortes implications écologiques et évolutives [10]. Notons que le cycle des saisons et le cycle lunaire y participent également (lire Focus Cycles de la lumière).
Le fractionnement temporel de l’environnement favorise la coexistence des espèces compétitrices et l’équilibre proie/prédateur. Si plusieurs espèces compétitrices n’ont pas accès à une même ressource en même temps, les confrontations directes entre ces espèces sont limitées. De même, si la ressource a le temps de se renouveler entre deux périodes de prédation, la ressource reste disponible pour plusieurs espèces. Ainsi, la coexistence entre compétiteurs est assurée. L’équilibre proie/prédateur est soumis à des fluctuations temporelles et prévisibles du risque de prédation. Les diverses espèces impliquées vont alors adopter un profil d’activité qui diminuera le risque de prédation tout en favorisant l’accès aux ressources de l’environnement [11]. Par exemple, les petits mammifères aux moyens de défense limités ont adopté au cours de l’évolution un profil d’activité nocturne qui limite le risque de prédation. Là aussi, la coexistence entre proies et prédateurs est assurée.
Le fractionnement temporel de l’environnement et l’adoption de profils d’activité différents ont imposé aux espèces différentes contraintes, anatomiques, physiologiques ou comportementales. Ainsi, les espèces diurnes utilisent la vision pour la communication et la prédation et leur rétine est riche en cônes. A l’inverse, les espèces nocturnes utilisent l’odorat, l’audition et le toucher pour la communication et la prédation. Leurs yeux possèdent de larges pupilles, parfois même une membrane capable de réfléchir la lumière (le tapetum lucidum) et leur rétine est riche en bâtonnets. Ainsi, en raison des différences d’intensité lumineuse entre le jour et la nuit, des adaptations complexes ont évolué et émergé pour accompagner les différents profils d’activité [11].
4.2. La lumière naturelle et les individus
La lumière et ses cycles, parce qu’ils procurent des informations non-visuelles et visuelles, sont indispensables aux organismes vivants. Pour une espèce donnée, les informations non-visuelles procurées par la lumière, telle que l’intensité lumineuse ou la photopériode, permettent aux individus de s’orienter dans leur milieu, de suivre un gradient de lumière et de synchroniser leurs rythmes biologiques pour anticiper les changements saisonniers de l’environnement. La capacité d’anticipation est essentielle pour tirer bénéfice des saisons favorables et se protéger des saisons défavorables. Pour anticiper les changements saisonniers de l’environnement, les organismes doivent mettre en place au moment opportun des caractères physiologiques et comportementaux, spécifiques et adaptés à la situation : migration, accumulation de réserves nutritionnelles, hibernation et reproduction. A cet effet, il leur est indispensable de synchroniser leurs rythmes biologiques avec un indicateur géophysique susceptible de servir de « calendrier ». C’est le rôle assuré par la lumière et ses cycles [1],[11].

Enfin, pour une espèce donnée, les informations visuelles procurées par la lumière, tels que les contrastes et les couleurs, permettent aux individus de détecter des objets sur le fond visuel, de naviguer dans leur milieu en relation avec des éléments inanimés ou vivants (Figure 7) et d’exprimer des caractères colorés qui servent au camouflage, au mimétisme ou sont associés au dimorphisme sexuel. Les informations visuelles procurées par la lumière sont donc indispensables pour la navigation, la recherche alimentaire, la prédation, la communication et la reproduction [1].
En conclusion, la lumière et ses cycles naturels représentent donc une importante force de sélection qui, au travers de l’évolution, a conditionné l’apparition de systèmes visuels et non visuels et d’horloges biologiques chez les organismes vivants. A l’échelle des individus, la lumière et ses cycles naturels ont permis aux organismes de développer une représentation spatiale et temporelle de leur environnement leur permettant de contrôler, d’exploiter et d’anticiper les changements du milieu. A l’échelle des écosystèmes, la lumière et ses cycles naturels assurent également le fractionnement des habitats dans le temps et dans l’espace, lequel est un facteur clé des équilibres inter- et intra-spécifiques.
Références et notes
Image de couverture : Photo © Klausi Shippe, Pixabay
[1] Nilsson DE (2009) The evolution of eyes and visually guided behaviour. Philosophical Transactions of the Royal Society of London: Biological Sciences 364, 2833–2847.
[2] Fernald R (2006) Casting a genetic light on the evolution of eyes. Science 313, 1914–1918.
[3] Arendt D (2003) Evolution of eyes and photoreceptor cell types. The International Journal of Developmental Biology 47, 563–571.
[4] le Tallec T (2013) Lumière, vision et horloge biologique. Espèces n°9, pp 12-21.
[5] Rodieck R (2003) La vision. Bruxelles, De Boeck Université.
[6] Koukkari WL & Sothern RB (2006) Introducing biological rhythms. New-York, Springer Verlag.
[7] Challet E (2007) Minireview: entrainment of the suprachiasmatic clockwork in diurnal and nocturnal mammals. Endocrinology 148, 5648–5655.
[8] Voigt R, Forsyth C & Keshavarzian A (2013) Circadian disruption: potential implications in inflammatory and metabolic diseases associated with Alcohol. Alcohol Research: Current Reviews 35, 87–96.
[9] Dibner C, Schibler U & Albrecht U (2010) The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annual Review of Physiology 72, 517–549.
[10] Kronfeld-Schor N & Dayan T (2003) Partitioning of time as an ecological resource. Annual Review of Ecology, Evolution, and Systematics 34, 153–181.
[11] Bradshaw WE & Holzapfel CM (2007) Evolution of animal photoperiodism. Annual Review of Ecology, Evolution, and Systematics 38, 1–25.
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Pour citer cet article : LE TALLEC Thomas (18 avril 2021), Lumière, vision et horloges biologiques, Encyclopédie de l’Environnement. Consulté le 30 juillet 2026 [en ligne ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/vivant/lumiere-naturelle-emergence-de-vie/.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.


