


Light, vision and biological clocks
PDF
Since life appeared on Earth, various selection forces have influenced the evolution of living organisms. Among these forces, light generated by our sun probably had the most significant influence. In particular, light and its natural cycles – daily and seasonal – have led living organisms to represent their environment in space and time. This has allowed them to adapt to this environment. How? What visual and non-visual photoreceptor systems have emerged in the animal kingdom? How have the benefits provided by these systems, vision and biological clocks, enabled organisms to control their environment and anticipate its changes?
Our star, the Sun, is 4.6 billion years old. It is a real nuclear reactor: every second, at its heart, 620 million tonnes of hydrogen are converted into 616 million tonnes of helium. The missing four million tonnes are converted into energy radiated outwards, i.e. photons, in other words light. Every second, after an 8-minute journey at a speed of 300,000 km per second, 6400 thousands billions photons per square millimeter enter the Earth’s atmosphere. A part of them is absorbed or reflected towards space by the atoms and molecules that compose it. The other part reaches the earth’s surface and illuminates our planet.
Since life appeared on Earth nearly 3.8 billion years ago, there have been 1,400 billion day/night cycles and 3.8 billion seasonal cycles (see Focus Light cycles and living organisms). Light has thus strongly influenced the evolution of the living world with the appearance of photosensitive systems, visual or non-visual, and biological clocks. In other words, light and its cycles have allowed living organisms to represent and adapt to space and time [1].
1. Evolutionary history of photoreception
1.1. Photoreceptor proteins
Different steps have marked the evolution of light-sensitive systems, from the appearance of the first photosensitive proteins to the current complex visual systems. An important class of photosensitive proteins is at the origin of photoreception: the opsin class. There are two super-families of them, the opsins of type I and II [2]. Type I opsins, also called microbial opsins, are found in the three domains of living organisms (Archae, Bacteria and Eukarya). They mainly regulate the orientation reactions of free organisms with respect to light, or photo-taxia, as well as the production of energy. When did these type I opsins appear? Their general distribution in all organisms suggests that their precursor already existed before the separation of the three domains of living organisms. This distant origin demonstrates the fundamental importance of light for life. Type II opsins, also known as animal opsins, are found only in higher eukaryotic organisms. They mainly regulate visual photoreception and biological clocks. The precursor of this type of opsin appeared 700 million years ago in primitive metazoans, the first mobile and heterotrophic multicellular organisms. The structure of these two types of opsins is similar, yet there is no phylogenetic link between these two super-families. On the contrary, they would have evolved independently of each other [2].
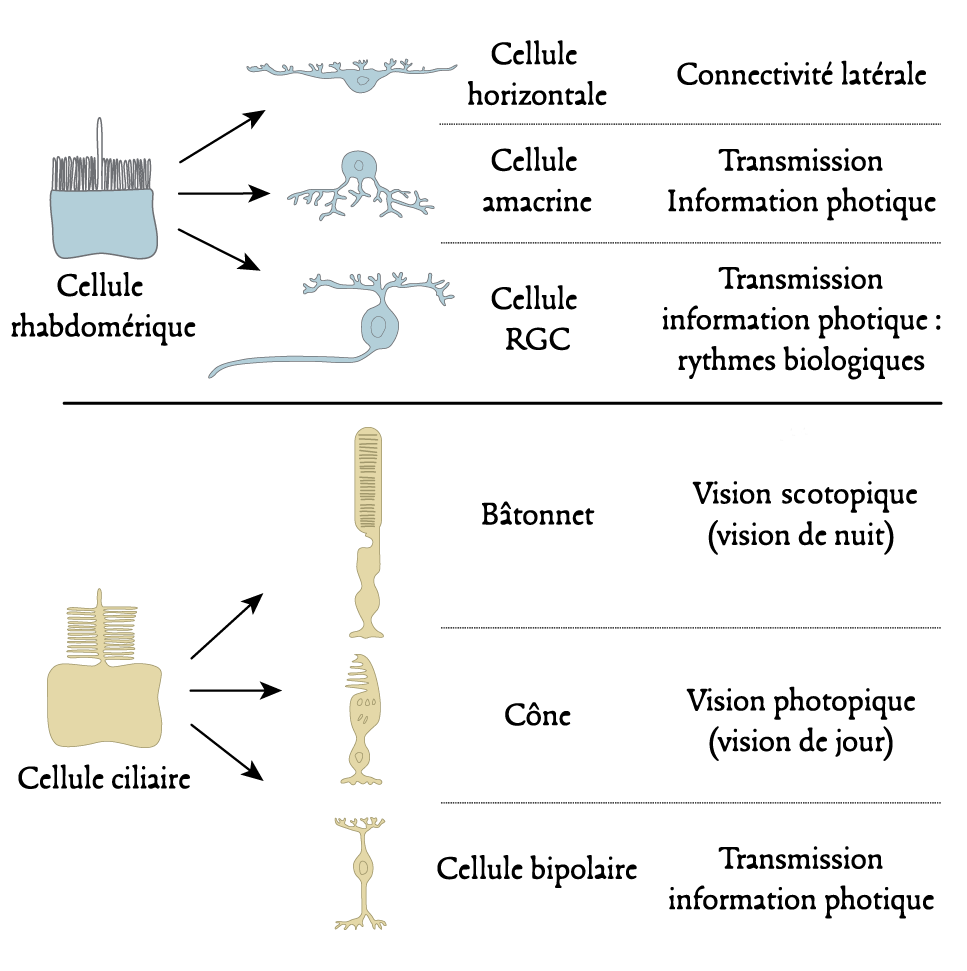
1.2. The photoreceptor cells
1.3. Non-visual and visual systems
Nowadays, most animals have complex non-visual and visual systems. Among the non-visual systems, we can distinguish various structures:
– an organ located inside the skull, in the shape of a small apple: the pineal organ; it is photosensitive in all vertebrates except mammals;
– an organ associated with the pineal organ, hence its name as a parapineal organ, intracranial in fish;
– a frontal organ or parietal eye in amphibians and reptiles;
– deep brain photoreceptors in all vertebrates except mammals;
– dermal photoreceptors, present on the skin in several vertebrate and invertebrate species, particularly amphibians and cephalopods.
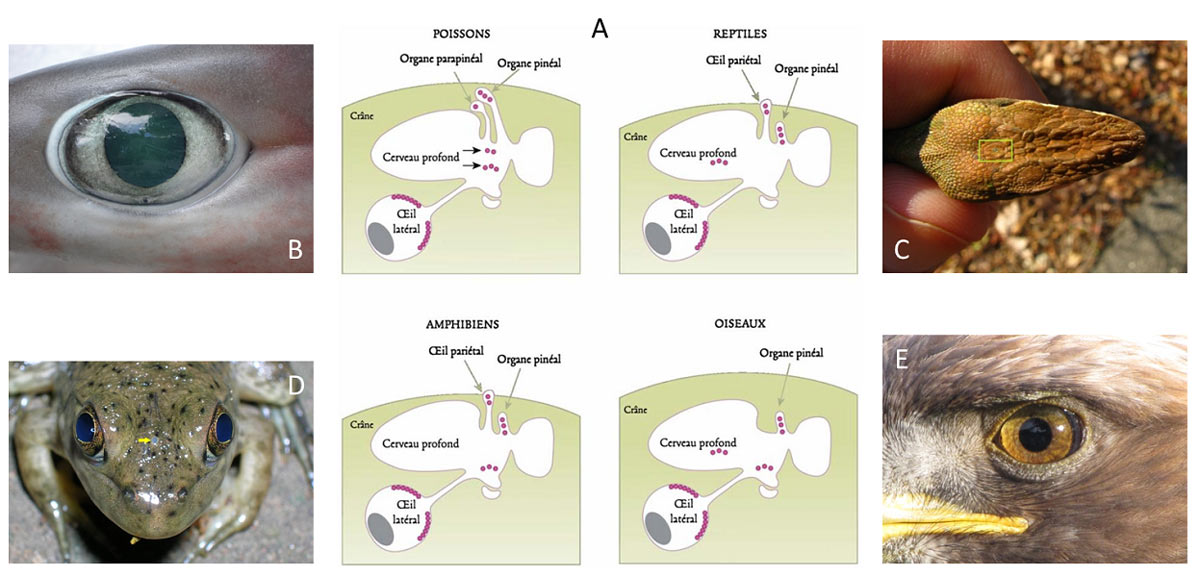
All these organs are involved in non-visual photoreception and play an important role in the regulation and control of biological clocks (Figure 2).
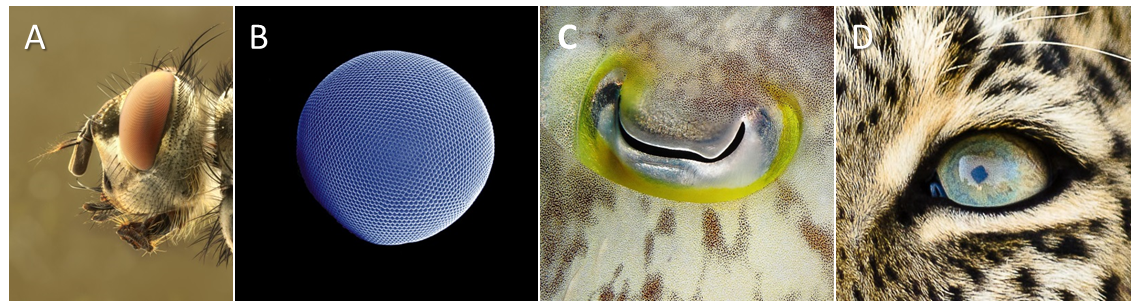
2. The vision
In all vertebrates, the lateral eyes have a common structure. It is similar to that of a single lens camera: the light passing through the pupil (lens) forms an inverted image on the retina (the sensitive surface) that lines the posterior surface of the eye. The retina is a tissue sheet composed of five types of connected neurons: visual photoreceptors, horizontal cells, bipolar cells, amacrine cells (without axons) and ganglion cells (Figure 4). A common distinction is between an inner and an outer part [5] (Figure 4).
There are two forms of visual photoreceptors, cones and rods. The former are sensitive to high light intensities and are therefore involved in so-called photopic day vision. The latter only react to low light intensities and are therefore involved in night vision, known as scotopic vision. The diversity of these photoreceptors is very high, both within the same species and between different species. However, each type of cone and rod contains a specific photoreceptor protein that is only sensitive to certain wavelengths of the electromagnetic spectrum. Each animal species therefore has a monochromatic or polychromatic vision that is specific to it (see below). Thus, fish, amphibians, reptiles and birds are, for the most part, tetrachromates: they process visual information using four types of photoreceptors. Mammals are generally dichromates, most likely because of their evolutionary history associated with nocturnal ecological niches. A number of primates, such as humans, are a notable exception, as they are trichromate [3]. This property, new to mammals, appeared following the duplication of a gene (see Focus Evolution of colour vision in mammals and primates).
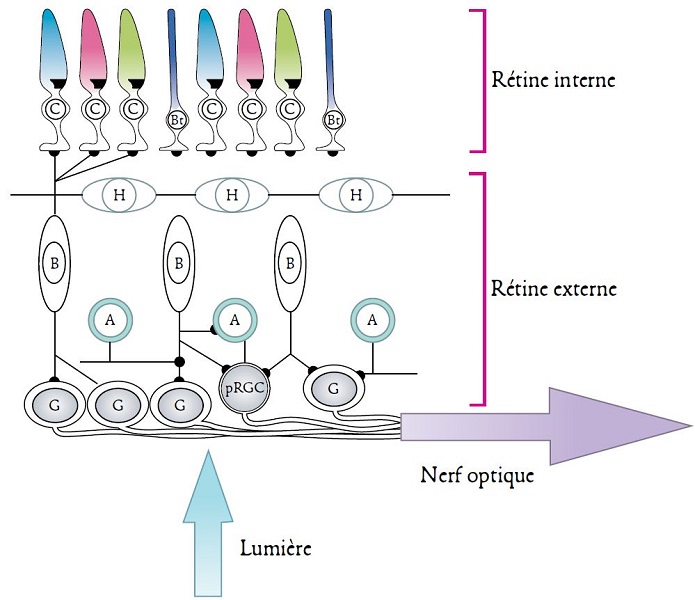
Thus, organisms are able to perceive the visual information emanating from their environment and can therefore make a spatial representation of it.
3. Clocks and biological rhythms
3.1. Biological rhythms and the day/night cycle
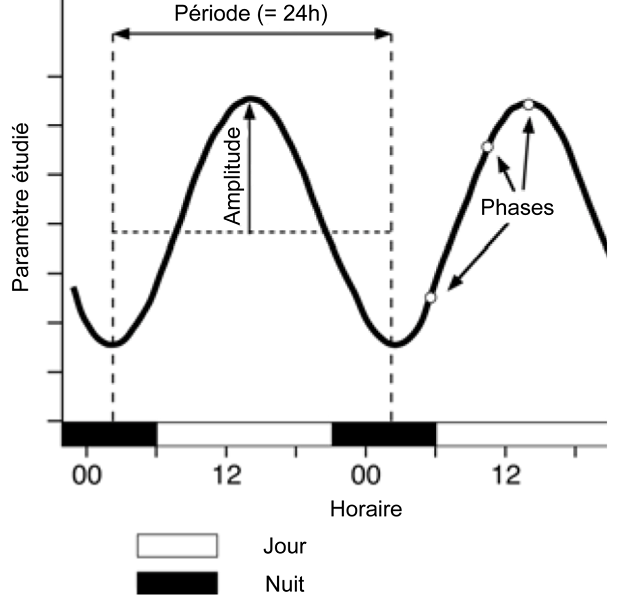
The term “biological rhythm” refers to a phenomenon that, within a living organism, is repeated over a period and amplitude that is constant over time. For a given species, these rhythms are genetically determined and generated by an internal clock that is active even in the absence of external stimuli. It is possible to distinguish three types of biological rhythms:
– circadian rhythms, also called daily rhythms, which have a period close to 24 hours;
– ultradian rhythms, which have a period of less than 24 hours;
– infradian rhythms, which have a period of more than 24 hours; the circadian rhythm, close to a terrestrial year, falls into this last category [6].
3.2. Central and peripheral clocks
In mammals, the main internal clock resides in the suprachiasmatic nucleus of the hypothalamus. This nucleus is composed of about 20,000 neurons capable of spontaneously and cyclically activating “clock” genes. Without external stimuli, and depending on the species, the expression of these genes oscillates over an endogenous period between 23h and 25h. Once activated, these clock genes in turn control the expression of other genes. In total, in mammals, nearly 20% of the genome is under the control of the main internal clock [7].
There are also secondary peripheral clocks. Indeed, clock genes are expressed in most peripheral tissues. However, without the main internal clock, the expression cycles of the clock genes differ from one peripheral tissue to another. The suprachiasmatic nucleus is therefore necessary to synchronize these secondary clocks.
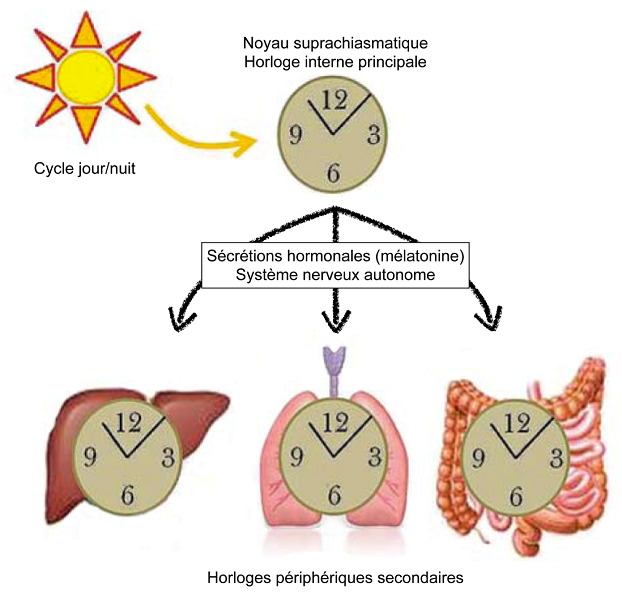
Once synchronized, the suprachiasmatic nucleus imposes its rhythm on brain structures and secondary peripheral clocks via hormonal secretions and activation of the autonomic nervous system. In particular, the suprachiasmatic nucleus modulates the activity of the pineal gland. During the day, it inhibits the activity of the cells of this gland: the pinealocytes. At night, he stimulates them. Pinealocytes produce and secrete the sleep hormone melatonin. Once secreted and released, melatonin binds to specific receptors located on the cell membrane of secondary peripheral clock cells, indicating to the body that it is dark. The suprachiasmatic nucleus thus imposes its rhythm on the whole organism [9] (Figure 6).
4. Natural light and its roles in life
4.1. Natural light and ecosystems
During a 24-hour day, the light intensity perceived by living organisms varies by several magnitudes. In other words, there is a specific light intensity for each hour of the day: the day/night cycle thus splits the environment. This is why, in animal species, there are different profiles of daily activities: daytime, nocturnal, twilight and cathedral activities (both day and night). This fractionation therefore has strong ecological and evolutionary implications [10]. It should be noted that the cycle of seasons and the lunar cycle also participate (see Focus Light cycles and living organisms).
The temporal fractionation of the environment favours the coexistence of competing species and the prey/predator balance. If several competing species do not have access to the same resource at the same time, direct confrontations between these species are limited. Similarly, if the resource has time to renew itself between two periods of predation, the resource remains available for several species. In this way, coexistence between competitors is ensured. The prey/predator balance is subject to temporal and predictable fluctuations in the risk of predation. The various species involved will then adopt an activity profile that will reduce the risk of predation while promoting access to environmental resources [11]. For example, small mammals with limited defences have adopted a nocturnal activity profile during evolution that limits the risk of predation. Here too, coexistence between prey and predators is ensured.
The temporal fractionation of the environment and the adoption of different activity profiles have imposed different anatomical, physiological or behavioural constraints on species. Thus, day species use vision for communication and predation and their retinas are rich in cones. In contrast, nocturnal species use smell, hearing and touch for communication and predation. Their eyes have large pupils, sometimes even a membrane capable of reflecting light (tapetum lucidum) and their retina is rich in rods. Thus, due to the differences in light intensity between day and night, complex adaptations have evolved and emerged to accompany the different activity profiles [11].
4.2. Natural light and individuals
Light and its cycles, because they provide non-visual and visual information, are essential to living organisms. For a given species, the non-visual information provided by light, such as light intensity or photoperiod, allows individuals to orient themselves in their environment, follow a light gradient and synchronize their biological rhythms to anticipate seasonal changes in the environment. Anticipation is essential to take advantage of favourable seasons and protect against unfavourable ones. To anticipate seasonal changes in the environment, organisms must put in place physiological and behavioural characteristics, specific and adapted to the situation: migration, accumulation of nutritional reserves, hibernation and reproduction. To this end, it is essential for them to synchronize their biological rhythms with a geophysical indicator that can serve as a “calendar”. This is the role played by light and its cycles [1],[11].

In conclusion, light and its natural cycles therefore represent an important selection force which, through evolution, has conditioned the appearance of visual and non-visual systems and biological clocks in living organisms. At the individual level, light and its natural cycles have enabled organisms to develop a spatial and temporal representation of their environment, enabling them to control, exploit and anticipate changes in the environment. At the ecosystem level, light and its natural cycles also ensure the fragmentation of habitats over time and space, which is a key factor in inter- and intra-specific balances.
References and notes
[1] Nilsson DE (2009) The evolution of eyes and visually guided behaviour. Philosophical Transactions of the Royal Society of London: Biological Sciences 364, 2833-2847.
[2] Fernald R (2006) Casting a genetic light on the evolution of eyes. Science 313, 1914-1918.
[3] Arendt D (2003) Evolution of eyes and photoreceptor cell types. The International Journal of Developmental Biology 47, 563-571.
[4] Le Tallec T. (2013) Lumière, vision et horloge biologique. Espèces 9, 12–21. (in french)
[5] Rodieck R (2003) La vision. Brussels, De Boeck University.
[6] Koukkari WL & Sothern RB (2006) Introducing biological rhythms. New York, Springer Verlag.
[7] Challet E (2007) Minireview: entrainment of the suprachiasmatic clockwork in diurnal and nocturnal mammals. Endocrinology 148, 5648-5655.
[8] Voigt R, Forsyth C & Keshavarzian A (2013) Circadian disruption: potential implications in inflammatory and metabolic diseases associated with Alcohol. Alcohol Research: Current Reviews 35, 87-96.
[9] Dibner C, Schibler U & Albrecht U (2010) The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annual Review of Physiology 72, 517-549.
[10] Kronfeld-Schor N & Dayan T (2003) Partitioning of time as an ecological resource. Annual Review of Ecology, Evolution, and Systematics 34, 153-181.
[11] Bradshaw WE & Holzapfel CM (2007) Evolution of animal photoperiodism. Annual Review of Ecology, Evolution, and Systematics 38, 1-25.
The Encyclopedia of the Environment by the Association des Encyclopédies de l'Environnement et de l'Énergie (www.a3e.fr), contractually linked to the University of Grenoble Alpes and Grenoble INP, and sponsored by the French Academy of Sciences.
To cite this article: LE TALLEC Thomas (January 5, 2025), Light, vision and biological clocks, Encyclopedia of the Environment, Accessed August 3, 2026 [online ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/en/life/light-vision-biological-clocks-2/.
The articles in the Encyclopedia of the Environment are made available under the terms of the Creative Commons BY-NC-SA license, which authorizes reproduction subject to: citing the source, not making commercial use of them, sharing identical initial conditions, reproducing at each reuse or distribution the mention of this Creative Commons BY-NC-SA license.


