


How do plants cope with alpine stress?
PDF
At high altitudes, plants can be subjected to strong constraints: high light and UV radiation, low CO2 concentration, large temperature variations… These parameters vary over the seasons but can also change significantly from one day to another as well as during the same day. These constraints modulate and sometimes disturb photosynthesis, which uses the energy provided by light and allows the growth and development of plants living in the alpine environment. In particular, excess light energy leads to the formation of toxic compounds, such as reactive oxygen species (ROS). To adapt to these difficult conditions, alpine plants developed various strategies: very small size, protective screen against UV radiation, protective anatomical structures, mechanisms to dissipate excess light energy, detoxification of reactive oxygen species, etc.
1. Vegetation in the alpine stages
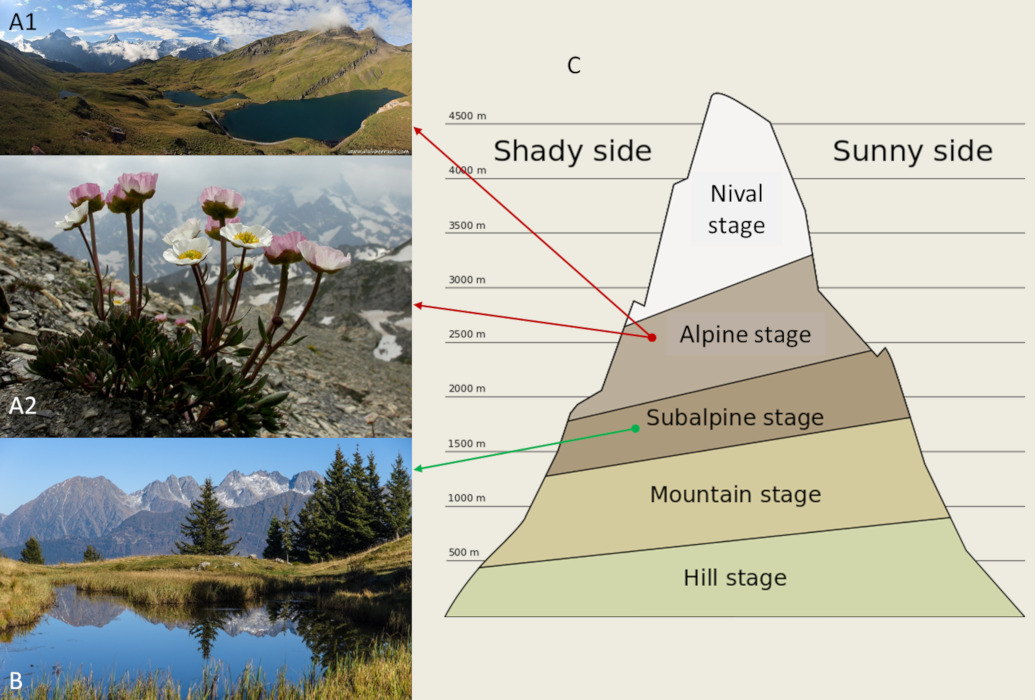
2. Climatic conditions and the growing season

Thus, perennial plants have the advantage of neo-forming only a few organs during the growing season whereas annual plants must form all their organs, which consumes a lot of energy for a limited time.

3. Photosynthesis
During the growing season, plants ensure their growth and development through photosynthesis. In this process, the necessary energy is provided by solar radiation, which allows the synthesis of organic matter from carbon dioxide taken from the atmosphere, and water and minerals taken from the soil. Because photosynthesis is affected by multiple environmental constraints such as those present at high altitude [5], it is likely that the maintenance of photosynthesis under stress conditions determines the survival of plants at high altitude. The main reactions of photosynthesis are summarized in Figure 4 and detailed in [6]. Figure 4 also describes the particularities implemented by alpine plants to protect this process.
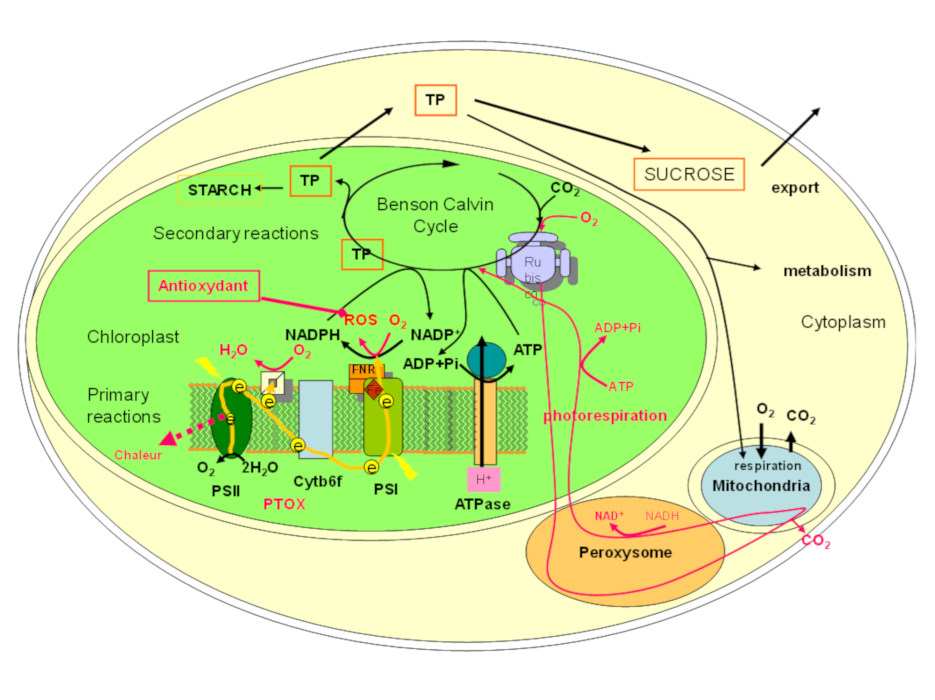
Photosynthesis takes place in the chloroplastsOrganites of the cytoplasm of photosynthetic eukaryotic cells (plants, algae). As a site of photosynthesis, chloroplasts produce O2 oxygen and play an essential role in the carbon cycle. They use light energy to fix CO2 and synthesize organic matter. They are thus responsible for the autotrophy of plants. Chloroplasts are the result of the endosymbiosis of a photosynthetic prokaryote (cyanobacterium type) within a eukaryotic cell, about 1.5 billion years ago. and can be divided into two phases.
- A primary phase takes place in the membrane of the thylakoids: the solar energy is absorbed by photosynthetic pigments (chlorophylls forming collecting antennas), then transferred to two reaction centres (photosystem I and II: PSI and PSII) whose coordinated activity results, via a succession of redox reactions, in the reduction of NADP+ to NADPHSigle for the reduced form of the coenzyme Nicotinamide Adenine Dinucleotide Phosphate (NADP). NADP is formed from Nicotinamide Adenine Dinucleotide (or NAD) by binding a phosphate to the 2′ hydroxyl group of the ribose associated with adenine. It exists in an oxidized form, called NADP+, and a reduced form, called NADPH. The NADPH is said to carry reductive power: used in catalyses carried out by oxidoreductases, it is capable of supplying energy during the transfer of their hydrogen atom, allowing the reduction reactions necessary for cellular functioning. by the PSI. The electrons involved come from the oxidation of water (2H2O → O2 + 4H+ + 4e–) at the PSII level. This primary phase also induces the establishment of a proton gradient on the thylakoid membrane which provides the energy necessary for the synthesis of ATPAbbreviation of adenosine triphosphate. A triphosphate nucleoside composed of adenine (nitrogen base), ribose (sugar with 5 carbon atoms) and three phosphate groups forming a triphosphate group. A compound that both donates and stores energy present in all living organisms. Also used as building materials for nucleic acid synthesis. (Figure 4).
- The second phase of photosynthesis takes place in the chloroplast stroma. It consumes the ATP and NADPH formed to fix atmospheric CO2 on a sugar di-phosphate (activity of RuBisCOAbbreviation for ribulose-1,5-bisphosphate carboxylase/oxygenase. It is the key enzyme for fixing CO2 carbon dioxide in plant biomass by initiating the Benson & Calvin cycle, using solar energy captured by chlorophyll during the photosynthesis process.), whose skeleton is formed by 5 carbon atoms (Ribulose bisphosphate or RuBP) and give sugar-phosphates with three carbon atoms (Triose-phosphate, in short TP) (Figure 4). This type of photosynthesis, which quickly leads to the formation of compounds with 3 carbons, is characteristic of C3 plants (C3 for 3 three carbons). The latter constitute the vast majority of alpine plants.
Trioses phosphate (TP), which contain assimilated CO2, can be used in chlorophyll-containg cells in several ways: (1) to regenerate the CO2 acceptor; (2) for the synthesis of glucose and starch in the chloroplast; (3) to be exported to other compartments of the cell where they provide energy and carbon chains for its maintenance; (4) for the synthesis of sucrose which is exported to other parts of the plant as an energy source and supplier of carbon skeletons feeding numerous biosyntheses [6]. Note also that oxygen can bind to RuBP: this oxygenation is at the origin of a metabolic pathway, photorespiration, which also consumes ATP and NADPH (Figure 4).
4. Importance of environmental factors in photosynthesis
4.1. Temperature
During photosynthesis, temperature mainly affects reactions allowing the binding of CO2 and O2 and the synthesis of sugars, but also the exchange of molecules between cellular compartments. The two main phases of photosynthesis and the transport processes involved are affected differently by temperature:
- Biophysical processes such as light absorption by chlorophyll pigments and the formation of NADPH and ATP are not or only slightly temperature dependent.
- The biochemical reactions of CO2 and O2 assimilation and sugar synthesis, as well as the exchange of molecules between cell compartments, are highly temperature dependent [7]. On average, a 10°C increase doubles the rate of biochemical reactions [6].
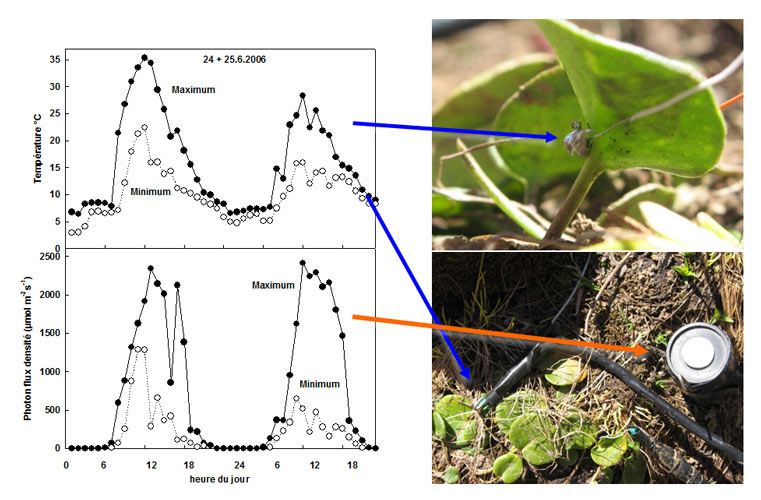
For example, the glacier buttercup, under light providing 2000 µmol photons m-2s-1, fixes about 15 µmol CO2 m-2s-1 at 23°C (see Figure 5 for the order of magnitude of the luminous flux), and only fixes half of it when it is at 10°C. In the first case the light is not saturated and the fixation of CO2 can eliminate the energy it provides, in the second case, however, the fixation of CO2 is already saturated under 500 µmol photons m-2 s-1 and therefore the light is largely excess [8].
4.2. Light
Like temperature, light can change suddenly by several orders of magnitude during the same day in alpine conditions (Figure 5). Under cloudy sky, the average light intensity is not very different between valleys and high altitude [3]. On the other hand, the maximum light intensity in clear weather is much higher at high altitude, exceeding the photosynthetic capacity of most plants.
Just like a drop in temperature, very strong light causes an excess of energy in the leaves that can damage photosystems [7]. In addition, there is more ultraviolet radiation (UVA and UVB) at higher altitudes [9], which can directly impact cellular structures: photosystems, but also DNA in the chloroplast, mitochondria and nucleus (see Cellular impact of solar UV). At the PSII level, manganese, which is involved in oxygen production (Figure 4), and plastoquinone, which transports electrons between the PSII and cytochrome b6/f (cytb6f), directly absorb UV radiations and in this way their function can be blocked [10].
A shade plant takes advantage of the moderate light intensities, which are common in the Alps, but is stressed if the light intensity is too high. On the other hand, a sun plant is more tolerant to high light intensities, but does not use weak light as well [11]. Finally, bright alpine plants such as Soldanella alpina, Homogyne alpina or Ranunculus glacialis (Figure 2) do not show any typical specific adaptation to low or high light [11].
4.3. Other factors
Other alpine environmental factors affect photosynthetic activity:
- The decrease in air pressure at high altitudes, for example, reduces the availability of CO2 [7].
- The wind – whose strength depends on the topography – increases the risk of plants drying out.
- By promoting rustle, the slope also reduces water retention by the soil [3].
However, in case of water stress, a plant reacts by closing its stomata (specialized cells controlling the gas exchange of the leaf): this limits the supply of CO2 and slows down photosynthesis. Here again, the biochemical reactions of photosynthesis cannot consume the excess light energy received by the plant.
In summary, climatic factors (such as light, temperature, humidity) vary strongly, rapidly and with large amplitudes in the mountains. If the majority of lowland plants are exposed to such variations, they do not survive, because photosynthesis is not able to use the excess energy [7]. This excess energy can be transferred to oxygen by forming reactive molecular oxygen species (ROS) (Figure 4 and [12]). ROS are potentially very destructive molecules; they can damage not only the photosynthetic apparatus and particularly photosystem II (photoinhibitionA process by which excess light decreases the speed of photosynthesis in organisms capable of performing it.), but also the entire cell (seeThe fixed life of plants and its constraints). In the end, the plant’s photosynthetic capacity may be inhibited by ROS, and its ability to repair damage is greatly reduced [11].
4.4. What are the constraints for photosynthesis of alpine plants?
The constraints imposed by the alpine environment can be illustrated, for example, in two extreme cases.
Some alpine plants such as Soldanella alpina, Geum montanum and Homogyne alpina keep some green leaves during the winter. Photosynthesis of these leaves begins immediately after snowmelt, under conditions that combine high light intensities with high thermal gradients [7]. These alpine species must take advantage of the short period after snowmelt for their photosynthesis, because they will be shaded by the other plants a few weeks later. Inactivation of photosynthesis in extreme climates can be lethal to these plants.
However, most alpine plants do not keep their leaves in winter and must quickly mobilize their reserves to form their first leaves and ensure their development cycle for a short favourable period. The case of Ranunculus glacialis (see Figure 2), which is only found above 2200 m, shows that this can be even more complex. Indeed, in this plant, it is calculated that the reserves mobilized for leaf formation represent about 30 days of photosynthesis, which can hardly be achieved during the growing season [3]. As Ranunculus glacialis flowers need two years to mature [12], this suggests that the energy received during a single growing season is not sufficient to complete its development cycle in one year. Ranunculus glacialis cannot therefore risk inactivation of photosynthesis. Dwarfing of several alpine species can also be considered as a strategy limiting the investment of energy in the formation of photosynthetic tissues.
5. Acclimation in alpine plants
Acclimation involves responses to minimize the amplitude of variations in the physical parameters of the environment at the plant level, before they exert their negative effects at the cellular and molecular level. When these responses are insufficient, acclimation is manifested through other protections to prevent the destruction of cells or their organelles.
Alpine plants often have a particular architecture in pillows (see Figure 3), rosettes or tussocks (see Inheritance or convergence). This type of morphology puts the plants in a favourable microclimate [3] with: (1) a decrease in light intensity and UV radiation aborbed by leaf tissue; (2) an increase in moisture at the level of leaves with wind protection, limiting drying out; (3) lower temperature variations than in the surrounding air. The acquisition of this type of morphology represents a first avoidance response. Of course, the growth of these plants is much slower than that of the plants that do not form these structures: this is the price to pay.
As described above, very large thermal variations combined with high luminosity can induce photoinhibition of photosystem II and the formation of ROS.
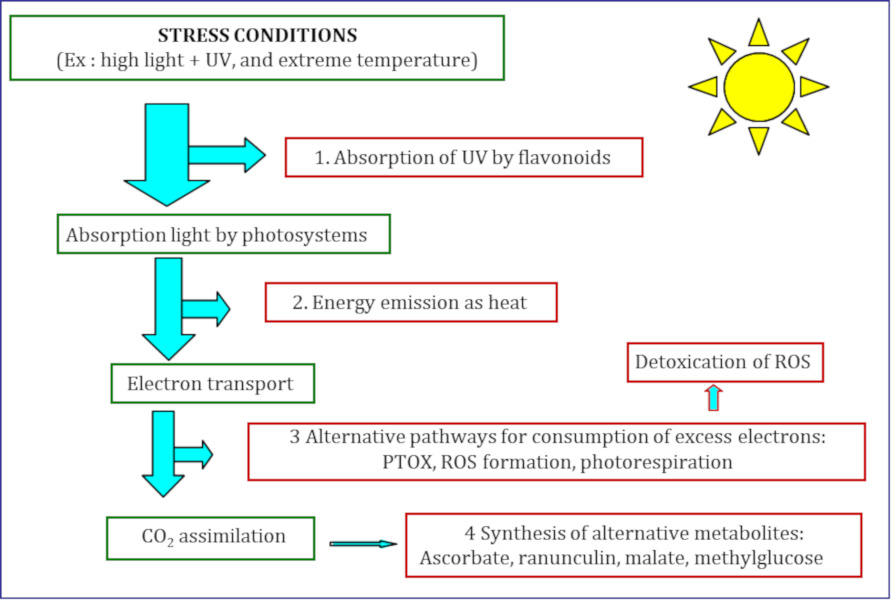
- Protection against UV radiation. At the epidermal level, some species synthesize flavonoidsSecondary metabolites of plants, all sharing the same basic structure. With several thousand compounds, flavonoids are the most important category of polyphenols and represent a gigantic family of antioxidants. Flavonoids are responsible for the brown, red and blue tones of flowers and fruits., pigments that absorb high-energy UV radiation. Thus, UV does not excite the underlying cells. This flavonoid screen thus protects the photosystems and prevents the destruction of the DNA of chlorophyll-containing parenchymesTissues composed of chlorophyll cells containing many chloroplasts. As the site of photosynthesis, they participate in nutritional functions. They represent an important part of the inside of the leaves.. In Soldanella alpina, for example, protection by high flavonoid contents is induced by strong light and increases with altitude [13].
- Re-emission of the excess energy absorbed. Light collecting antennas change conformation in illuminated plants; at the same time, some of the excited chlorophylls pass their energy to carotenoids (zeaxanthin for example). This allows some of the light energy to be dissipated as heat (Figure 4). In this way, the amount of energy arriving at the reaction centres is controlled. Under excess light, the thermally dissipated light fraction is higher than that directed towards the reaction centres. Thermal dissipation is a quick way to remove the excited states of chlorophyll. However, while in some alpine species, such as Soldanella alpina and Geum montanum, this avoidance mechanism is very effective, in others, such as Ranunculus glacialis, it is not very effective and other mechanisms are activated (see below) [7].
- Energy transfer to alternative acceptors via an electron flow. In chloroplast, the alternative oxidase (called PTOXEnzyme of the membranes of the chloroplast thylacoids, which catalyzes the oxidation of the plastoquinone pool, hence its name alternative oxidase (PTOX).), connected between the two photosystems (Figure 4), accepts electrons provided by the PSII and transfers them to molecular oxygen, thus creating water. This enzyme can thus reduce electronic pressure and thus the formation of ROS under excess light. Compared to lowland plants, the content of PTOX in alpine plants is high and its concentration increases with altitude in Geum montanum [7],[8].
- Maintaining photosynthetic metabolism at low temperatures. If the CO2 supply limits the synthesis of sugars in the second phase of photosynthesis, Rubisco catalyzes the binding of molecular oxygen to RuBP. This is the first step in photorespiration (Figure 4). Photorespiration consumes energy, in the form of (a) carbon (RuBP), (b) electrons produced during the first phase of photosynthesis and (c) ATP. The activity of photorespiration, usually negligible at low temperatures, becomes important when the latter exceeds 20°C. In Ranunculus glacialis, which does not tolerate heat, photorespiration is already active when the temperature is low [7],[8].
- Synthesis of specific metabolites. Some alpine plants synthesize metabolites that are generally absent or present in small quantities in lowland plants. The function of these metabolites is often not understood. But their synthesis consumes part of the energy captured by the photosynthetic apparatus and can thus help to protect it. Thus, Ranunculus glacialis accumulates significant amounts of ranunculinGlucoside produced by plants such as ranunculus (Ranunculaceae). A highly unstable molecule, it is hydrolyzed into an irritating lactone: proto-anemonin and malateMalic acid salt, a dicarboxylic acid widely used in the plant kingdom and naturally present in fruit, which contributes to its pleasant taste. Malate is an intermediate of the Krebs cycle, one of the major metabolic pathways of cellular respiration in almost all living beings, and is involved in the Benson & Calvin cycle, which is part of photosynthesis. Used as a food additive, under number E296 [14]. Geum montanum contains high concentrations of methylglucoseMonosaccharide (glucose) with a methyl group (CH3)., while Soldanella alpina contains record amounts of ascorbate (vitamin C) in its leaves [15]. Ascorbate is part of the antioxidant system that protects the plant from ROS.
- Detoxification of highly reactive forms of oxygen (ROS). Environmental constraints increase the formation of ROS, especially during the early stages of photosynthesis. In Figure 4, ROS formation is indicated at the PSI level: it is the major pathway for the formation of ROS in light. To degrade ROS to oxygen, plants use a set of enzymatic reactions with associated metabolites. In Soldanella alpina leaves, the antioxidant system is based on the presence of vitamin C and vitamin E, as well as enzymes involved in redox processes [7]. In Potentilla saundersiana, the importance of the antioxidant system of the leaves increases with altitude [16].
In short, it is a set of protection mechanisms that allow alpine plants not only to survive, but also to develop in a habitat that becomes more and more restrictive with altitude. These mechanisms complement and add up to facilitate the life of alpine plants (Figure 6).
References and notes
Cover image. Alpine linaria (Linaria alpina) in the Galibier schists (2600 m). [Source: Photo © Serge Aubert/SAJF]
[1] Ozenda P., La végétation de la chaîne alpine dans l’espace montagnard européen, Masson, 1985
[2] Fischesser B., La vie de la montagne, Éditions de La Martinière, Paris, 1998
[3] Körner C., Alpine plant life, Springer Verlag, Berlin Heidelberg, 1999
[4] Larcher W., Kainmüller C., Wagner J., Survival types of high mountain plants under extreme temperatures, Flora, 205:3-18, 2010
[5] Lütz C., Plants in Alpine Regions, Cell Physiology of Adaptation and Survival Strategies, Springer, Wien New York, 75-97, 2012
[6] Raven P., Evert R., Eichhorn S., Biology of Plants, Sixth Edition, W.H. Freemann and Company/Worth Publishers, 1986
[7] Streb P., Cornic G., Photosynthesis and Antioxidative Protection in Alpine Herbs, In Lütz C. (Ed) Plants in Alpine Regions: Cell Physiology of Adaptation and Survival Strategies, Springer Wien New York, pp 75-97, 2012
[8] Streb P., Josse E.-M., Gallouët E., Baptist F., Kuntz M., Cornic G. (2005) Evidence for alternative electron sinks to photosynthetic carbon assimilation in the high mountain plant species Ranunculus glacialis. Plant, Cell Environment 28:1123-1135
[9] Barry R., Mountain weather and climate, Cambridge University Press, Third Edition, 2008
[10] Teramura A., Ziska L., Ultraviolet-B radiation and photosynthesis, In “Advances in Photosynthesis Vol. 5: Photosynthesis and the Environment“. Ed. N.R. Baker, pp 435-450, Kluwer Academic Publishers, Dordrecht, 1996
[11] Walters R., Towards an understanding of photosynthetic acclimation, Journal of Experimental Botany 56:435-447, 2005
[12] Ort D., Baker N., A photoprotective role of O2 as an alternative electron sink in photosynthesis? Current Opinion in Plant Biology 5:193-198, 2002
[13] Laureau C., Meyer S., Baudin X., Huignard C., Streb P. (2015) In vivo epidermal UV-A absorbance is induced by sunlight and protects Soldanella alpina leaves from photoinhibition. Functional Plant Biology, 42:599-608
[14] Streb P., Aubert S., Gout E., Bligny R. (2003) Reversibility of cold- and light-stress tolerance and accompanying changes of metabolite and antioxidant levels in the two high mountain plant species Soldanella alpina and Ranunculus glacialis. J. Exp. Bot. 54:405-418
[15] Bligny R., Aubert S., Specifities of metabolite profiles in alpine plants. In Lütz C. (Ed) Plants in Alpine Regions: Cell Physiology of Adaptation and Survival Strategies, Springer, Wien & New York, pp 99-120, 2012
[16] Lan Ma, Xudong Sun, Xiangxiang Kong, Jose Valero Galvan, Xiong Li, Shihai Yang, Yunqiang Yang, Yongping Yang, Xiangyang Hu, Physiological, biochemical and proteomics analysis reveals the adaptation strategies of the alpine plant Potentilla saundersiana at altitude gradient of the Northwestern Tibetan Plateau, Journal of Proteomics, 112:63-82, 2015
The Encyclopedia of the Environment by the Association des Encyclopédies de l'Environnement et de l'Énergie (www.a3e.fr), contractually linked to the University of Grenoble Alpes and Grenoble INP, and sponsored by the French Academy of Sciences.
To cite this article: STREB Peter, CORNIC Gabriel, BLIGNY† Richard (January 5, 2025), How do plants cope with alpine stress?, Encyclopedia of the Environment, Accessed July 9, 2026 [online ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/en/life/how-do-plants-cope-with-alpine-stress-2/.
The articles in the Encyclopedia of the Environment are made available under the terms of the Creative Commons BY-NC-SA license, which authorizes reproduction subject to: citing the source, not making commercial use of them, sharing identical initial conditions, reproducing at each reuse or distribution the mention of this Creative Commons BY-NC-SA license.