


Plant resistance to extreme temperatures
PDFPlants have a reduced mobility and have no other choice than to face the variations of the physical conditions of their environment. In order to adjust and adapt to new conditions they must perceive the changes and translate them into signals capable of being directed to a target that will perform the operations that will allow them to maintain their activity. The response of plants to their environment therefore involves receptors and signalling molecules.
- The targets can be (1) the nuclear, chloroplast or mitochondrial genome, (2) transcription factors, or (3) transcription factor activators. Syntheses are then initiated or modulated, or even stopped. The response often affects both metabolism and morphogenesis.
- Note that a signal from a modification perceived only at one point of the plant (on one of its leaves, for example) can be transmitted to the whole plant, and provoke a global adaptive response that is called systemic response.
It is difficult to identify a single thermal receptor that would link the plant to its environment: the plant probably has several at key points of metabolism. However, it seems necessary that the signals they produce interact to produce the appropriate global response. Finally, it should be noted that the diversity of systems that “perceive” temperature changes can contribute to the diversity of the plant world.
Research using the possibilities of molecular genetics has developed in the last two decades. It has considerably deepened our knowledge of the response to extreme temperatures. But many points remain to be clarified. This focus presents a few avenues that will give the reader an idea of the complexity of the problems posed.
1. Hardening and light: the “phytochrome” thermometer
The interactions between light and cold hardening have long been highlighted (see Effect of temperature on photosynthesis). In some cases, cold hardening can even be obtained at ordinary temperature by modulating the length and spectral composition of the photoperiod. However, cold hardening is still necessary to obtain a complete hardening
Phytochromes (P) are a family of chromoproteins that allow the plant to perceive the photoperiod length and spectral composition in the red (Figure 1). They are well known to be involved in the processes of morphogenesis and flowering.
- Their inactive form (PR) preferentially absorbs red light (R) to give an active form PFR preferentially absorbing dark red (FR).
- This reaction is reversible: PFR absorbing RS gives again the inactive form (PR). The PFR/PR ratio in the plant exerts control over many plant responses to light (morphogenesis, flowering etc.) by controlling signalling pathways leading to modulation of gene activity.
- But the switch from PFR (active form) to PR (inactive form) also occurs independently of light. This reaction is strongly influenced by temperature. It can therefore modulate the PFR/PR ratio, and consequently the activity of phytochromes, independently of the characteristics of the light environment.
- It constitutes a thermometer able to appreciate a difference of 1°C, at least. [1]
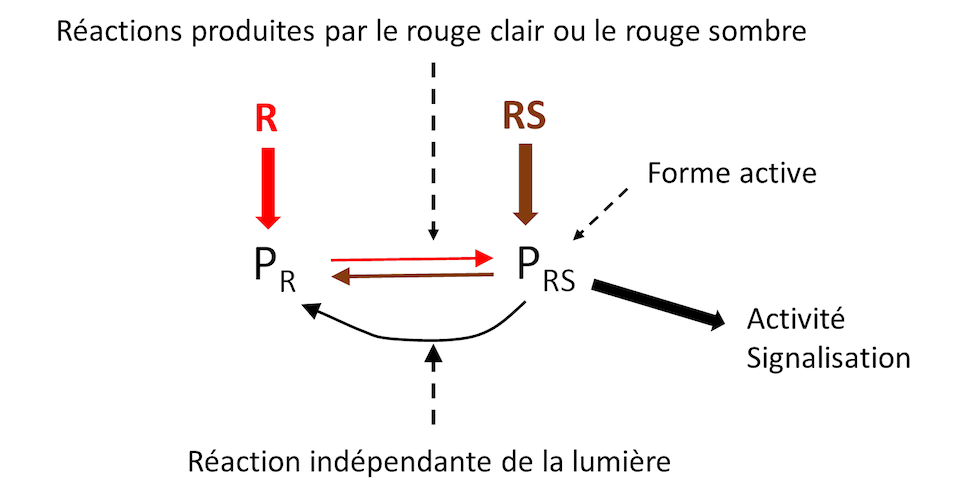
PFR, transferred into the cell nucleus, modulates cellular responses caused by low or high temperatures. This regulation occurs at the level of transcription factors that control, among other things, several aspects of morphogenesis [2].
2. Membrane fluidity: its relationship with related proteins
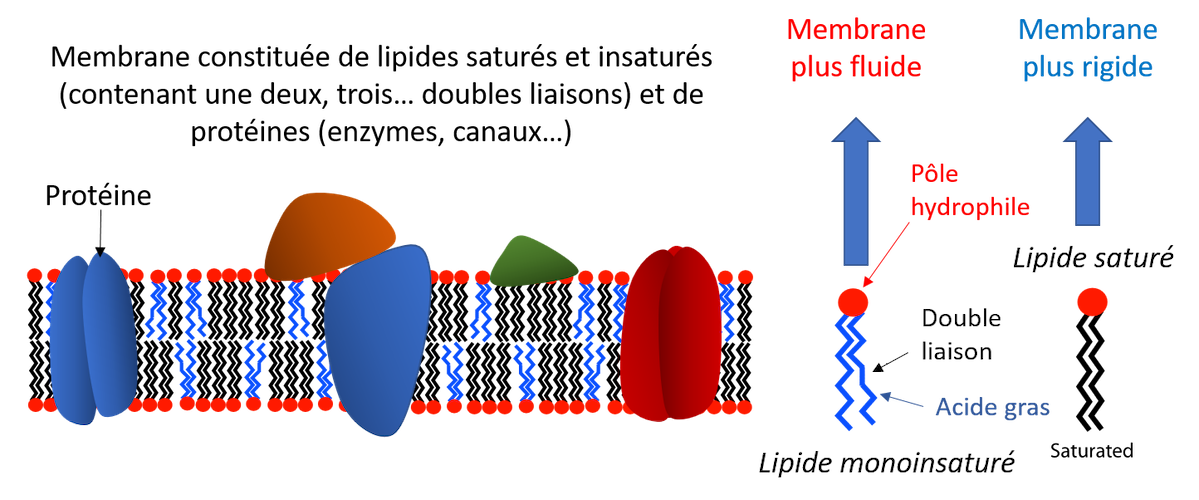
Biological membranes, lipid bilayers, are composed of a mixture of unsaturated fatty acids (with one or more double bonds), which are fluid, and saturated fatty acids (do not have double bonds), which are comparatively more rigid.
Their fluidity increases with temperature, affecting not only their permeability but also the activity of the proteins included in them or interacting with them.
This modification of activity can thus be the cause, by successive cascades, of a modulation of the activity of genes.
However, fluidity tends to be maintained, despite temperature variations, by a rapid modulation of the ratio between saturated and unsaturated fatty acids [3], with an influence on the activity of the proteins included in it.
3. Isoprene synthesis
Isoprene is emitted into the atmosphere by many woody species (such as oaks, eucalyptus, conifers) and herbaceous species. Several species of mosses (such as Sphagnum) and ferns also produce isoprene. However, many plants do not emit isoprene: this is the case, for example, of the bean, although other papilionaceous plants produce it.
Isoprene protects the photosynthetic apparatus [4] against thermal shock. This protection is also effective in plants that emit little or no isoprene, placed in an atmosphere that contains levels compatible with those measured near the leaves of emitting plants.
Isoprene is thought to contribute to the rapid stabilization of membranes, but also to the regulation of the quantity of active O2 species. Finally, it acts as a signalling molecule by acting on all the systems involved in growth and in responses to stress.
4. The production of active oxygen species (ROS)
The production of active oxygen species is common in plants subjected to all kinds of constraints (physical or biological). It occurs, at both low and high temperatures, mainly in chloroplasts, mitochondria and peroxisomes.
For example, active oxygen species (ROS) [5] are continuously produced during photosynthesis. Indeed, as the absorption of light by the collecting antennae and its transfer to the reaction centers of the photosystems are insensitive to temperature, any decrease in the assimilation of CO2 produced by extreme temperatures (low and high) will result in excess light energy in the photosystems (Read How do plants cope with alpine stress?)
- Part of this energy is evacuated in the form of heat from the antennae;
- Another part contributes to the formation of ROS, which can damage the photosynthetic apparatus or trigger acclimatisation responses.
ROS are formed at other stages in the functioning of the system of transduction of light energy into chemical energy. They trigger adaptive responses by interacting with the metabolic and signalling networks already at work.
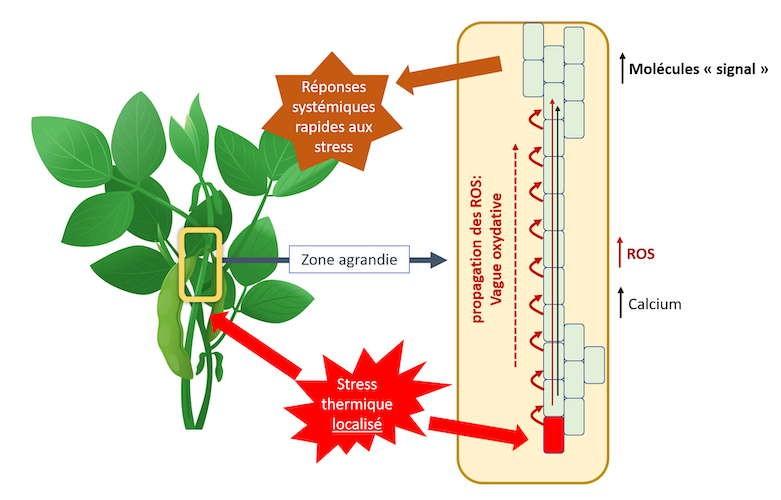
They also participate, probably with calcium, in systemic responses. Indeed, it has been shown that an oxidative wave arises at the point of the plant subjected to an increase in temperature and spreads to the entire plant. It originates from the production of ROS at the point affected by the stress (Figure 3). This propagation occurs from cell to cell in two ways:
- By a symplasmic pathway, i.e. from cytoplasm to cytoplasm via plasmodesmata and channels, such as aquaporins;
- By an apoplastic pathway, located between the skeletal wall and the plasmalemma. This last pathway involves an NADPH oxidase located in the plasmalemma whose activation requires Ca++. A ” calcium wave” accompanies the oxidative wave.
Notes and references
Cover image. [Source: © G.Cornic]
[1] Jung, J-H., Domijan, M., Klose, C., Biswas, S.,Ezer, D.,Gao, M., Khattak, A.K., Box, M.S.,Charoensawan, V., Cortijo, S., Kumar, M., Grant, A., Locke, J.C.W., Schäfer, E. , Jaeger, K.E., Wigge, P.A. (2016) Phytochromes function as thermosensors in Arabidopsis. Science 354, 886-889
[2] Catalá, R., Medina, J., Salinas, J. (2011) Integration of low temperature and light signaling during cold acclimation response in Arabidopsis. Proceedings of the National Academy of Sciences 108, 39, 16475-16480
[3] Sung DY, Kaplan F, Lee KJ, Guy CL (2003) Acquired tolerance to temperature extremes. Trends in Plant Science 8, 179-187
[4] Sharkey TD, Chen X, Yeh S (2001) Isoprene increases thermotolerance of fosmidomycin-fed leaves. Plant Physiology 125, 2001-2006
[5] Reactive Oxygen Species = ROS. ROS are radicals that have a single electron on their peripheral layer, a characteristic that allows them to interact easily with their environment.
[6] Suzuki N, Devireddy AR, Inupakutika MA, Baxter A, Mille G, Song L, Shulaev E, Rajeev K. Azad, Shulaev V, Mittler R (2015) Ultra-fast alterations in mRNA levels uncover multiple players in light stress acclimation in plants. The Plant Journal 84, 760-772