Terrestrial plants are regularly subjected to strong temperature variations. These variations can reach an amplitude of 40°C or even more, both in polar regions and in hot desert areas. Being rooted, they have reduced mobility and must cope with changes in their environment. The assimilation of CO2 by plants via photosynthesis is the gateway to carbon in the biosphere. What is the thermal amplitude that allows it to function? How does photosynthesis reacts to rapid and slow temperature variations? What is the diversity of responses? What are the physiological processes that limit it? Crucial questions tare o be considered in the context of global warming.
The current increase in greenhouse gas emissions will cause an increase in atmospheric temperature of 2 to 3°C in the next 50 years (see A carbon cycle disrupted by human activities). At the same time, heat waves and extreme heat periods will be more frequent and of longer duration [1]. Agricultural production and the functioning of forests will therefore be greatly affected. Models based on large-scale observations indicate that, in the absence of agronomic adaptation, the decrease in crop yields can reach 17% for each 1°C increase in the temperature of the growing season [2].
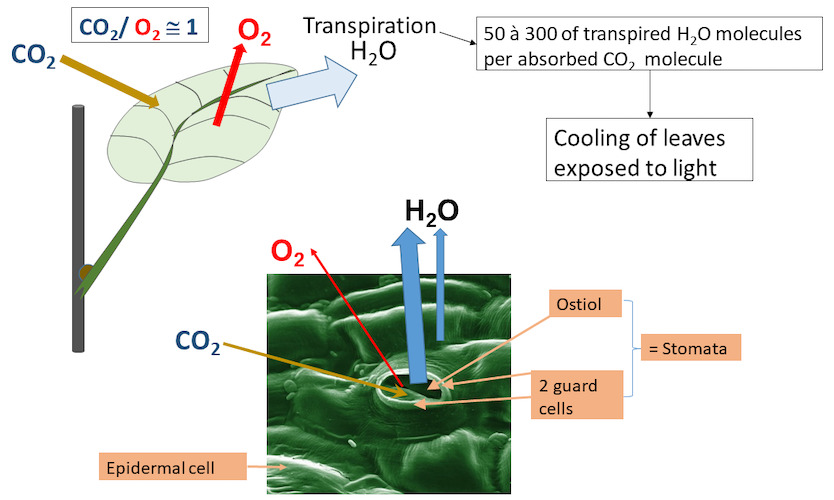
The production of higher plants depends in particular (but not only [3]) on leaf photosynthesis (see Shedding light on photosynthesis & The The path of carbon in photosynthesis). CO2 enters the leaf where its reduction in the chloroplasts is accompanied by O2 production. Its entry is almost exclusively through the stomata (Figure 1). For each molecule of CO2 absorbed, 50 to 300 molecules of water are transpired from the leaves, depending on the plant. This water allows, among other things, the cooling of the leaf (see Focus Leaf transpiration and heat protection).
Figure 1. During photosynthesis, CO2 is absorbed and O2 is released mainly through the stomatal opening (ostiole). The water vapour (transpiration of the leaf) passes mainly through the ostiole but also through the epidermis. Transpiration allows the leaf to cool down in the light. [Source: Author’s diagram]The leaf is a converter of solar energy into chemical energy and, like any energy converter, requires a permanent cooling system.
The climate changes that are currently occurring make it necessary to understand the effects of temperature on photosynthesis.
2. The thermal optimum of photosynthesis
2.1. Diagram of the thermal response
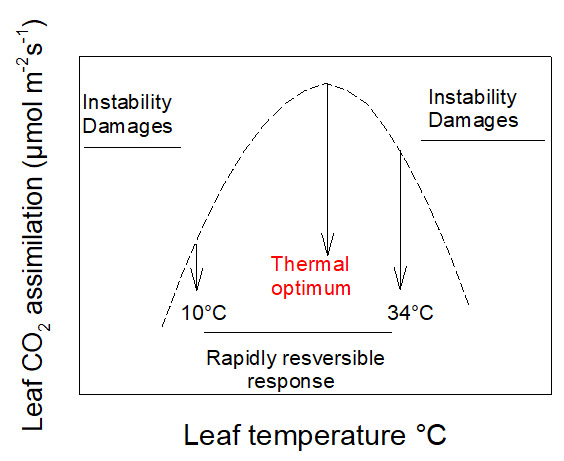
Photosynthetic CO2 uptake varies with temperature. In most cases its response to temperature is rapidly reversible between about 10 and 34°C. In this range of temperatures it presents a maximum value: a thermal optimum.
Figure 2. Diagram of the variation of CO2 assimilation by an intact leaf. It highlights the temperature range in which variations are generally rapidly reversible. [Source: Author’s diagram]Below 10°C and above 34°C plants start to set up protective mechanisms. For these extreme values, CO2 assimilation is often unstable and can be cancelled more or less quickly: the leaf is then under stress (Figure 2).
2.2. A thermal optimum based on the average temperature of the environment
Plants in cold environments or with a cold growing season have a higher photosynthesis at low temperatures. Plants in warm environments, or growing during the warm season, have a higher photosynthesis at high temperatures.
Figure 3. Deschampsia antarctica is one of two flowering plants found in Antarctica. It is often subjected to negative temperatures. The snow that frequently covers it protects it from extreme temperatures. [Source: Lomvi2, CC BY-SA 3.0, via Wikimedia Commons]For example, the thermal optimum for CO2assimilation [4] in Deschampsia antarctica (Figure 3) and Colobanthus quitensis, the only two Antarctic flowering plants, is between 8 and 15°C, while it is around 45°C in Tridestomia oblongifolia, a warm desert plant from Central America. The latter species probably holds the world record for flowering plants in this respect.
2.3. Acclimatization to the thermal conditions of the environment
Figure 4. Variations in CO2 assimilation as a function of leaf temperature, in a plant grown at 10°C (red) or 25°C. Measurements made on Pea, under a light close to saturation. CO2 content in ambient air: 390 ppm. [Source: Author’s diagram]Differences in the thermal response of photosynthesis are also found in individuals of the same species growing at different temperatures. Figure 4 shows CO2 assimilation in pea grown at 10 or 25°C.
In the first case (cultivation at 10°C) the thermal optimum is about 16°C, while it is higher than 25°C in the second (cultivation at 25°C). At low temperatures, CO2 assimilation is higher in plants grown at 10°C.
In this case the adjustment to cool conditions is a gain for the plant.
2.4. Acclimatization can be rapid
Figure 5. Remth (Hammada scoparia), a characteristic plant of the Wadi Rum desert (Jordan). [Source: Ji-Elle, CC BY-SA 4.0, via Wikimedia Commons]For example, the photosynthesis of Hammada scoparia, a bush in the deserts of the Middle East (Negev, Wadi Rum) follows the seasonal variations in temperature: its thermal optimum varies from 29°C in early spring to 41°C in summer and then to 28°C in autumn
In general, these changes can be measured in both growing and mature leaves, with the response being of greater amplitude in growing leaves.
2.5. Heat-sensitive versus cold-sensitive species
Warm acclimation of cool-adapted species (or ecotypes [6]) occurs with an increase in thermal optimum but a general decrease in photosynthesis.
This is, for example, the case of Atriplex sabulosa. One can then wonder about the interest of this change. The opposite may be true for plants strictly adapted to warm conditions, such as Tridestomia oblongifolia. Figure 7 illustrates the case of Atripex lentiformis, [7] a perennial leafy plant, which occurs in California in both Death Valley and in cool, wet coastal habitats:
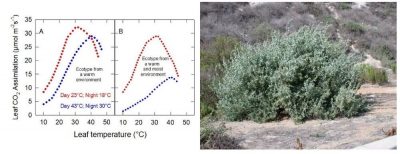
The assimilation of the desert ecotype (Figure 7A) and the coastal ecotype (Figure 7B) show almost the same response to temperature when grown under 23°C during the day and 18°C at night (in red in Figure 7).
Figure 7. Variations in CO2 assimilation of Atriplex lentiformis ecotypes from a warm environment (A) and a cool, moist environment (B). [Source: Author’s diagram, after Pearcy (1977)] Right, Atripex lentiformis (salt bush) [Source: Forest & Kim Starr, CC BY 3.0, via Wikimedia Commons]Under the alternating 43°C day and 30°C night (blue in Figure 7), only the desert ecotype shows plasticity, maintaining high CO2 assimilation under these new conditions. The activity of the coastal ecotype is low at all temperatures. Only the displacement of the thermal optimum remains of its acclimatization capabilities [8].
C4 plants, of which there are traces only from the end of the Tertiary Era, constitute only 5% of the species. They tend to colonize hot and dry environments (or seasons) (See Restoring savannas and tropical herbaceous ecosystems). Maize and sugarcane are examples.
On average, the thermal optimum of C4 plants is located at higher temperatures than that of C3 plants.
However, C3 plants are the most plastic. In fact, their thermal optimum varies from around 7 to 35 ° C, while that of C4 plants oscillates, with a few exceptions, between 30 and 40 ° C. In addition, when the temperature is below 20 ° C, the photosynthesis of C4 plants is on average lower than that of C3 plants.
3. CO2 assimilation results from the interaction of processes whose response to temperature is different
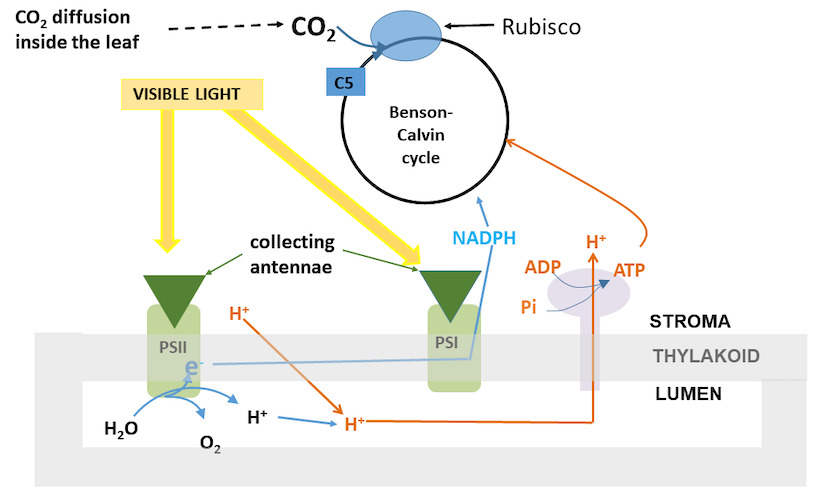
The absorption of light at the collecting antennae (Figure 9) and the transfer of its energy to the PSII reaction centres are not temperature sensitive. Are temperature sensitive:
The diffusion of CO2 from the ambient air to the chloroplasts: its speed increases with temperature.
Figure 9. Diagram of the interacting processes during photosynthetic CO2 fixation (case of a C3 plant). PSI and PSII: respectively photosystem I and II. They are included in the thylakoid membrane, which is made up of two lipid layers forming “sacs” in the chloroplast. The interior of the thylakoid is the lumen. RubisCO: enzyme that catalyzes the fixation of CO2 on a sugar with 5 carbon atoms (Ribulose 1,5-bisphosphate: C5). Benson-Calvin cycle: allows the regeneration of C5, and at the same time gives the plant the necessary carbon. ATP is synthesized when protons from the lumen return to the stroma through an ATPase using inorganic phosphate, Pi. The lumen protons have two origins: (1) oxidation of water in the lumen by PSII which also provides electrons, e- and (2) operation of a proton pump in the thylakoid that passes protons from the stroma into the lumen. [Source: Author’s diagram]The regeneration of RuBP occurs via the operation of the Benson-Calvin cycle (This is the “biochemistry” of the process) which uses reducing power (in the form of NADPH) provided by electron transfer to function. The necessary ATP is synthesized when protons accumulated in the lumen pass into the stroma through an ATPase (Figure 9). The formation of reducing power and the synthesis of ATP have a thermal sensitivity close to that of electron transfer.
4. What are the processes at work in setting the thermal optimum for CO2 assimilation in C3 and C4 plants?
4.1. Photosystem activity and the resulting electron transfer are not involved
Measured in vitro on isolated thylakoids (see legend Figure 9), in the presence of artificial acceptors, electron transfer increases with temperature and shows a clear thermal optimum. It is located around 30°C and corresponds to that of CO2 assimilation when the latter is saturating[9]. The activity of PSII has a thermal optimum identical to that of the electron transfer chain.
PSI activity is not inhibited at high temperatures (above 30°C, up to 45°C) where it remains stable or even increases: it is the activity of PSII that limits the activity of the electron chain.
Moreover, PSII is very sensitive to high temperatures which damage the protein complex that allows the oxidation of water (see Figure 9).
The thermal response of electron transfer is similar in C3 and C4 plants. However, there are organizational differences between these two types of plants (see The path of carbon in photosynthesis).
The supply of energy cannot therefore explain the differences in thermal optimum. It is the way in which the energy produced is used that makes the difference.
4.2. An answer? Comparison of the effect of atmospheric O2 on CO2 assimilation of C3 and C4 plants
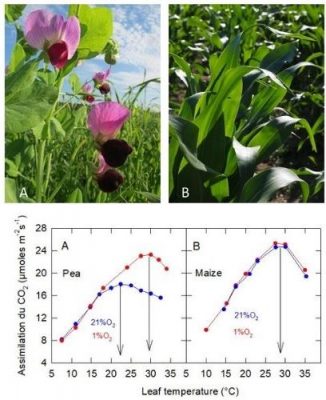
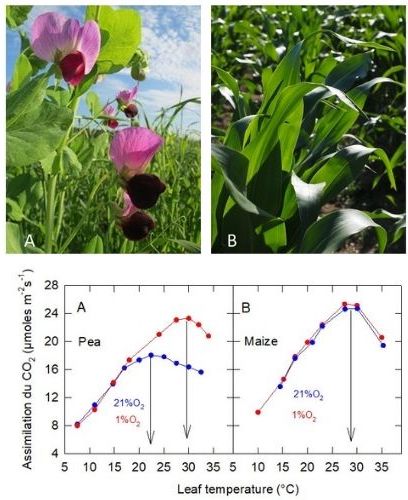
In normal air[10], 21% O2 (+ N2) + 360 ppm CO2: the thermal optimum is 27°C in Maize (C4 plant), while it is only 22°C in Pea (C3 plant) (Figure 10): the thermal optimum of the C4 plant is higher than that of the C3 plant (see also section 2.6).
In an oxygen-deficient atmosphere, 1% O2 (+ N2) + 360 ppm CO2: the CO2uptake of Maize is not affected, while that of Pea is stimulated above about 17°C, with a shift in its thermal optimum to near that of Maize.
In C3 plants, atmospheric oxygen inhibits CO2 uptake when the leaf temperature is sufficiently high, whereas it has no effect (or negligible effect) in C3 plants
Figure 10. Variation of CO2 assimilation measured in leaves of Pea (A; Pisum sativum) and Maize (B; Zea mays) as a function of leaf temperature. The plants were grown in natural light at a temperature of 20 ± 2°C. [Source: Author’s diagram – royalty-free image / Pixabay]Note that the variation in electron transfer estimated in vivo, by measuring chlorophyll fluorescence emission as a function of temperature, is very similar in 1% and 21% O2 in Pea: the variation in thermal optimum is therefore not due to a change in photochemistry.
4.3. Rubisco properties explain the difference in response
Case of C3 plants
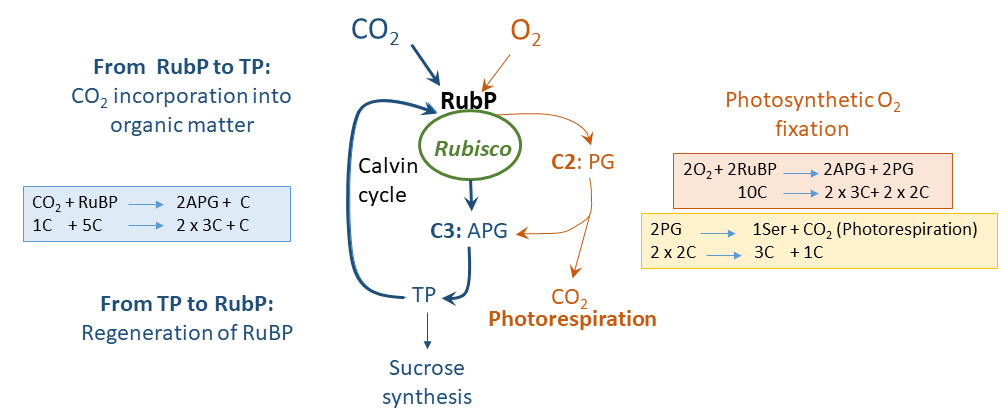
CO2 and O2 compete to occupy the active sites of Rubisco: This enzyme has a carboxylase function and an oxygenase function. CO2 enters the Benson-Calvin cycle and the photosynthetic fixation of O2is at the origin of a metabolic pathway responsible for photorespiration (Figure 11; see also The path of carbon in photosynthesis).
CO2 occupies a high number of active sites on the Rubisco when the O2 content of the ambient air is low (1% for example) or that of CO2 is high.
O2 is mainly fixed if its content increasesor if that of CO2 decreases (the latter then releases active sites which are then occupied by O2).
In normal air, there are two reasons why O2 fixation increases (and consequently CO2 fixation decreases) when the temperature increases[11].
The affinity of Rubisco forCO2decreases more than that for O2; a factor that favours the assimilation of O2.
The water solubility coefficient of CO2 decreases more than that of O2, leading to a more rapid decrease in the amount of CO2 than O2 in the chloroplast; this is a factor that favours O2 fixation.
In an O2-poor atmosphere (Figure 10), competition between O2 and CO2is very reduced. Energy is then used mainly for CO2 assimilation, which increases in value until around 30°C and then decreases as the energy supply decreases (see section 4.1).
In normal air, the effect of O2 on photosynthetic CO2 fixation (Figure 11) is very low (or even nil) when the temperature is low: competition on the carboxylation sites is in favour of CO2.
Figure 11. Schematic of CO2 and O2 fixation on RuBP (Ribulose 1,5-bisphosphate) in a C3 plant. APG: 3-phosphoglyceric acid, 3 C compound; TP: Trioses phosphate. The carbon leaves the Calvin cycle to feed the synthesis of sucrose. PG: phosphoglycolate, 2C compound. Two PGs give a serine (Ser) containing 3C with the production of CO2 from photorespiration. C = Carbon atom. Source: Author’s diagram]
On the other hand, when the temperature increases, the competition on these sites favours the fixation of O2which then consumes an increasing part of the energy produced by the activity of the photosystems. This energy is therefore no longer available for CO2 fixation, which reaches its maximum value around 22°C.
Case of C4 plants.
CO2 is concentrated at the Rubisco by a mechanism that is insensitive to oxygen. Its content can reach 800 to 2000 ppm depending on the plant in C4: that is to say contents from 2 to 5 times higher than its current atmospheric content.
Under these conditions, photosynthetic O2 fixation is weak or even non-existent because the active sites of the Rubisco are all occupied by CO2. The energy supplied by the activity of the photosystems is therefore used only in the fixation of CO2 when the leaf temperature increases, explaining the higher thermal optimum in this type of plant.
C4 plants evolved from C3 plants during the global decrease in atmospheric CO2content at the end of the Tertiary Era [12].
This decrease would then have “released” the oxygenase function of the Rubisco of C3 plants, resulting in a loss of fixed carbon via photorespiration.
The establishment of a CO2 concentration mechanism is an advantage because it prevents this carbon loss. We currently find species that are “intermediates” between C3 and C4.
5. The thermal optimum of C3 photosynthesis is modulated by certain environmental parameters
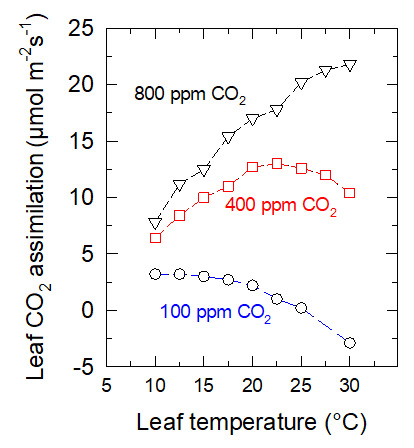
5.1. The CO2content in the atmosphere
The thermal optimum increases with increasing ambient CO2 content. In the case shown in Figure 12, it increases from about 10°C when the content is 100 ppm to more than 30°C when it is 800 ppm.
Figure 13. Human activities lead to an increase in carbon dioxide in the atmosphere. Its content went from 320 to 415 ppm in the space of 50 years. This increase has consequences on the temperature of the atmosphere and the activity of the vegetation. [Source : Royalty-free image / Pixabay] In a world with steadily increasingatmospheric CO2 (Figure 13), the thermal optimum of C3 plants is expected to increase. This does not mean, however, that plant production will then be higher (see note 3 section 1): episodes of high heat will, like droughts, certainly be more frequent.
5.2. Lack of water
The photosynthetic apparatus is resistant to drought. It retains all its capacity to absorb CO2 on the Rubisco, and to produce energy until the leaves have lost about 30% of their water [13].
CO2 uptake decreases in this range of water loss, because the stomata close (see Focus Leaf transpiration and heat protection). This closure slows down the entry of CO2into the leaf and consequently leads to a decrease of the CO2 content in the mesophyll.
However, the O2 content in the chloroplasts remains high. Indeed, its content in the atmosphere (21% or 210,000 ppm) is, compared to that of CO2 (@ 400 ppm), very high and in any case sufficient for a very substantial quantity to pass through the epidermis even when the stomata are closed.
The competition between CO2 and O2 for the occupation of the active sites of the Rubisco is thus in favour of O2.
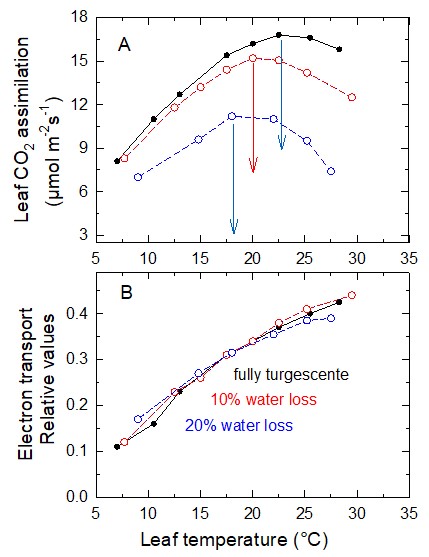
Figure 14. A, Variations in CO2 assimilation as a function of leaf temperature. Leaves with different amounts of water loss found in air with an ambient CO2 content of 400 ppm. B, The electron transfer rate estimated on the same leaves by measuring the chlorophyll fluorescence emission. [Source: Author’s diagram, after Cornic et al. ref. 14]Therefore, the thermal optimum for photosynthesis must lower in C3 plants that dry out.
This is shown in Figure 14A, in which the thermal optimum drops from about 23°C, in a Pea leaf at maximum turgor, to 17°C when it has lost 20% of its water.
Electron transfer in the thylakoid membrane is not affected by water loss in the range shown (Figure 14B). When water loss is 20%, the energy produced by photosystem activity is primarily used to bind atmospheric oxygen to RuBP [14], resulting in increased photorespiration.
6. Why, from its thermal optimum, CO2 assimilation decreases as temperature decreases or increases?
6.1. When the temperature lowers
Several reasons probably all contribute, to varying degrees, to this decrease :
The rate of RuBP turnover decreases: there is a slowdown in the activity of some enzymes controlling this turnover, notably that of a Fructose 1,6-bisphosphate (see Figures 9 and 11).
Sequestration of phosphorylated compounds in chloroplasts. The triose phosphate is no longer (or less) exported when sucrose synthesis is inhibited. The inorganic phosphate in the chloroplast is no longer renewed leading to a decrease in ATP synthesis.
Inhibition of the electron transfer chain (see section 4.1), resulting in reduced energy production (reducing power and ATP).
In C4 plants it is the activity of the Rubisco that appears to be preponderant, although the cold sensitivity of enzymes involved in CO2 accumulation at the Rubisco is well known.
6.2. As the temperature increases
In C3 plants the increase in photorespiration decreases the fraction of electrons produced by PSII and used to assimilate CO2. However, other factors are at play since CO2 assimilation measured (1) in an atmosphere with little or no photorespiration (ambient O2 content of 1%), and (2) measured in a normal atmosphere in a C4 plant decreases in both cases (Figure 10).
Several reasons can be given:
The slowing down of PSII activity leading to that of the electron transfer chain from PSII to PSI.
To perform its role Rubisco must be activated by an enzyme called Rubisco activase, the activity of which decreases when the temperature is higher than about 33°C (incidentally, high-temperature resistant activases appear in some plants subjected to periods of high heat [15]). However, since activase must itself be activated by an electron transfer-dependent process, it cannot be ruled out that the latter is also involved in limiting [15].
The “catalytic misfiring” of Rubisco increases with temperature and increasing amounts of an inhibitor of the enzyme (Xylulose-1,4-bisphosphate), which is structurally close to RuBP (see Figures 9 and 11), are synthesized.
In C4 plants (case of Maize) the activation and activity of enzymes that participate in the CO2 concentration system at the Rubisco are not very sensitive to high temperatures. The same reasons as above may explain the decrease in CO2 assimilation when the temperature increases beyond that of the thermal optimum.
7. Hardening after plant exposure to cool (≤ about 10°C) and high (≥ about 37°C) temperatures
Maintaining plants at cool or high temperatures causes, along with the changes in photosynthesis described above, increase in their resistance to otherwise lethal temperatures(frost and high temperature). This is hardening.
In this process, temperature and light interact and the metabolic changes induced are sometimes very rapid (from minutes to hours).
Thus, cold hardening can be achieved at ordinary temperature by modulating the length of the light period or its spectral composition in the red[16]. However, cold is still required to achieve full hardening. Also the lack of light in the cold prevents hardening to varying degrees.
At elevated temperatures : the transmitted signals activate the synthesis of chaperone proteins (HSPs: Heat Schock Proteins) that repair denaturing proteins, also prevent their coagulation or even help mark them for degradation.
At cool temperatures: the synthesis of chaperone proteins is also activated. It is accompanied by (i) the synthesis of “antifreeze” proteins that interfere with ice crystal formation and (ii) an increase in sugar synthesis tending to increase osmotic pressure in the cells.
Note that the signaling pathways and their interactions inducing the genome response are only partially known. The references given in “Learn More” and an attached Focus allow for further exploration of this evolving point.
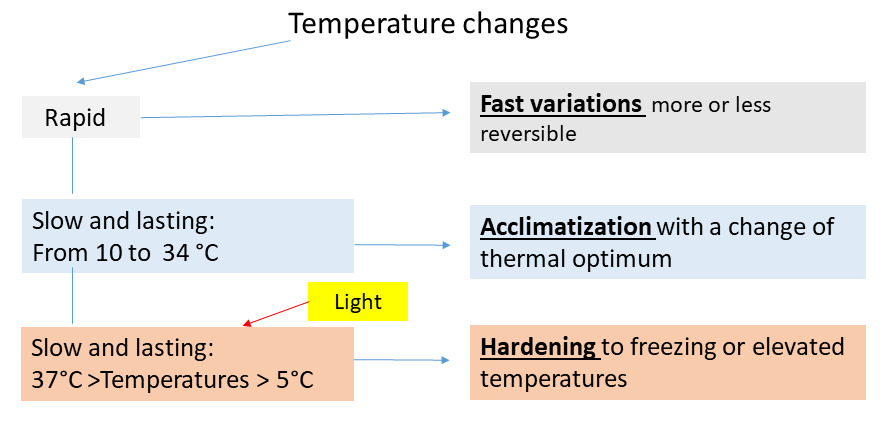
8. Effects of temperature on photosynthesis: summary diagram
The summary diagram (Figure 15) classifies the effects of temperature on photosynthesis according to the speed of temperature change and the extent of its variation. Note that hardening allows leaf maintenance in perennial leaf plants and therefore minimizes energy loss under extreme temperature conditions.
Figure 15. Scheme classifying the effects of temperature on photosynthesis. [Source: Author’s diagram]The rapidity of current climate change makes it necessary to delve deeper into the responses of plants to their environment: the hope is to be able to maintain sufficient primary production to keep the biosphere functioning.
9. Messages to remember
The uptake of CO2 by a leaf has a thermal optimum close to the average temperature of its growth environment.
This thermal optimum can change rapidly when the conditions of the environment are durably modified: this is a process acclimatization.
This thermal optimum is on average less in C3 plants than in C4 plants: this is mainly due to photosynthetic fixation of atmospheric O2 via Rubisco activity in C3 plants.
This optimum depends on the CO2 content of the ambient air in C3 plants: at high content it becomes identical to that in C4 plants.
This optimum depends on the hydration state of the leaf.
Subjected to cool or hot temperatures plants bring into play processeshardening to otherwise lethal temperatures. These processes involve protein syntheses and changes in the fluidity of chloroplast and cell membranes.
Notes and references
Cover image. Sunset over the Sonora Arizona desert. [Source: royalty free / Pixabay]
[1] Meehl GA, Stocker TF, Collins WD, Riedlingstein P, Gaye AT, Gregory JM, Kitoh A, Knutti R, Murphy JM, Noda A & Raper SCB (2007). Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press
[2] Yamori W, Hikosaka K & Way DA. (2014). Temperature response of photosynthesis in C3, C4, and CAM plants: temperature acclimation and temperature adaptation. Photosynthesis Res. 119, 101- 117.
[3] For example, when growing plants are subjected to drought, the amount of carbon they assimilate decreases initially because leaf growth is inhibited. The mechanisms for CO2 fixation in the leaf are not then inhibited. Boyer JS (1970) Plant Physiol.46, 233-235
[4] The values of thermal optima given here, are from measurements made in “normal air”, containing 21% O2 and about 400 ppm CO2. When this is not the case the O2 and CO2 contents are shown. The CO2 uptake in air containing 21% O2 is saturated from about 1200 ppm CO2 when light is close to saturation. The evaporative power of the air is also regulated in most cases during the measurements. It is estimated by the saturation deficit of the partial pressure of water vapor in the ambient air around the leaves.
[5] Plants from the same individual by vegetative reproduction. They are genetically identical.
[6] Ecotype: Plants of the same species from different environments, which, grown from seed to flower under identical conditions show different physiological characteristics.
[7] It fetches water from as far as the water table, hence its name of phreatophyte plant.
[8] Pearcy RW (1971). Acclimation of photosynthetic and respiratory CO2 exchange to growth temperature in Atriplex lentiJormis (Torr.) Wats. Plant Physiol. 59, 795-799
[9] Yamasaki T, Yamakawa T, Yamane Y, koike H, Satoh K & Katoh S. (2002) Temperature acclimation of photosynthesis and related changes in photosystem II electron transport in winter wheat. Plant Physiol. 128 1087-1097.
[11] Jordan DB & Ogren WL (1984). The CO2/O2 specificity of ribulose 1,5-bisphosphate carboxylase/oxygenase. Dependence on ribulose bisphosphate concentration, pH and temperature. Planta161, 308-313
[12] Ehleringer JR, Sage RF, Flanagan LB & Pearcy RW (1991). Climate change and the evolution of C4 photosynthesis. Trends in Ecology and Evolution 6, 95-99
[13] Cornic G & Massacci A (1996). Leaf photosynthesis under drought stress. In Advances in Photosynthesis (vol 5) Photosynthesis and the environment, 347-366. Neil R Baker (ed.) Kluwer Academic publishers Dordrecht.
[14] Cornic G, Badeck F-W, Ghashghaie J & Manuel N (1999). Effect of temperature on net CO2 uptake, stomatal conductance for CO2 and quantum yield of photosystem II photochemistry of dehydrated pea leaves. In Sanchez Dias M, Irigoyen JJ, Aguirreolea J & Pithan K (eds) Crop development for cool and wet regions of Europe. European community. ISBN 92-828-6947-4.
[15] Crafts-Brandner SJ, van de Loo FJ & Salvucci ME (1997). The two forms of ribulose-1,5-bisphosphate carboxylase/oxygenase activase differ in sensitivity to elevated temperature. Plant Physiol. 114, 439-444.
[16] Puhakainen T, Li C, Boije-Malm M, Kangasjärvi J, Heino P & Palva ET. (2004). Short-day potentiation of low temperature-induced gene expression of a C-repeat-binding factor-controlled gene during cold acclimation in Silver Birch. Plant Physiol.136, 4299-4307
The Encyclopedia of the Environment by the Association des Encyclopédies de l'Environnement et de l'Énergie (www.a3e.fr), contractually linked to the University of Grenoble Alpes and Grenoble INP, and sponsored by the French Academy of Sciences.
The articles in the Encyclopedia of the Environment are made available under the terms of the Creative Commons BY-NC-SA license, which authorizes reproduction subject to: citing the source, not making commercial use of them, sharing identical initial conditions, reproducing at each reuse or distribution the mention of this Creative Commons BY-NC-SA license.
Global climate change is a major concern. It is based on the scientific community’s statement…
Serge PLANTON, General engineer of bridges, waters and forests, former climatologist researcher at Météo-France, CNRM (Centre National de Recherches Météorologiques).