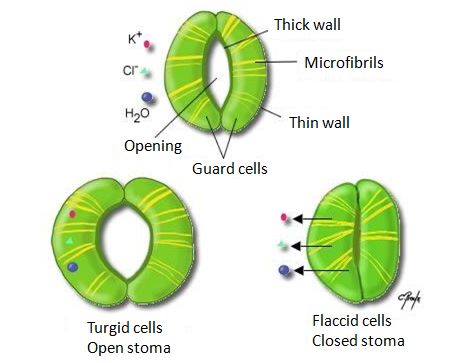
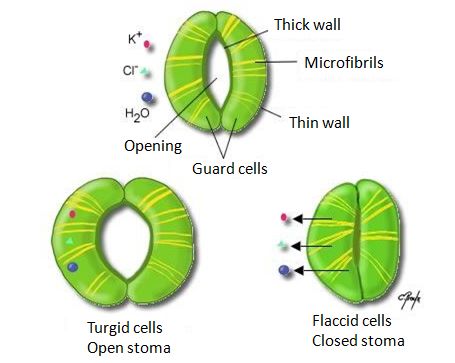
图6. 树木中水分根据水势递减而流动的循环图。[图片来源:作者自制]( Mesophyll cells 叶肉细胞,Xylem 木质部,Stoma 气孔,Transpiration draws water from the leaf 叶片通过蒸腾作用丧失水分,Cohension and adhesion draw water up the xylem 在木质部内,水分子的内聚力和粘附力促使其上行,Root hairs 根毛,Soil particle 土壤颗粒,Water molecule 水分子,Negative water potential draws water into the root 负水势使得水被吸入根中,Low 低,High 高,Atmosphere 大气,Leaf at top of tree 树顶端的叶片,Stem 茎干,Root cells 根细胞)土壤水势略为负(-0.05MPa或-0.5bar),因为地表以下的水,含有低浓度的溶解性矿盐。
[3] Dufrêne E, Pontailler J-Y and Saugier B, 1993. A branch bag technique for simultaneous CO2 enrichment and assimilation measurements on beech (Fagus sylvatica L.) Plant, Cell and Environment, 16, 1131-1138
Michel BAKALOWICZ, 巴卡洛维茨·米歇尔,前法国国家科学研究中心(CNRS)研究员, 前蒙彼利埃水文科学区域水与环境中心(CREEN)副主任,黎巴嫩贝鲁特高级工程学院退休教授,蒙彼利埃水文科学自由研究员。
The tireless-quest for water by plants
How does water reach the leaves of large trees? What tricks do plants develop to cope with the lack of water? How does vegetation modify the climate? Life was born in water, and land plants have had to adapt to temporary water shortages. The roots allow them to draw water from the soil, and the leaves, through their transpiration, exert a suction effect that draws this water through the stems. The water is therefore under tension or negative pressure in the stems. When this tension becomes too strong due to a lack of water, it causes cavitation with a gas bubble in the vessels which cease to be conductive. To avoid this, transpiration is regulated by the stomata, which close. The stomatal closure limits the entry of CO2 into the leaf and consequently photosynthesis and production.
This statement is trivial for anyone who has a few plants at home: they must be watered regularly, some very often, others less. All living things are made up of water-filled cells, and plants generally contain between 70 and 95% water. As always, there are a few exceptions: the wood of trees contains 30-70%, and seeds can dry out almost completely, but still be able to germinate.
1.2. Plants also need air, and oscillate between hunger and thirst
When the plant lacks water, it closes stomata to limit water loss, but this also stops CO2 uptake, leading to a risk of starvation. The plant must therefore constantly adjust the opening of its stomata to regulate its water loss while ensuring its subsistence. Stomata have existed since at least 400 Ma (millions of years), and plants have optimized their ability to exchange with the outside air since 325 Ma.
1.3. Is the water in plants under pressure or tension?
This water pressure exists permanently in plant cells and allows their growth: it pushes their walls, makes the cells swell and increase their volume and the size of plant organs.
But when trees have leaves, it is not possible to collect sap, because during the day the tree transpires and the water in the vessels is under tension, a negative pressure caused by the evaporation of water from the leaves. This is the most common situation for a plant, as we will clarify.
1.4. How does water circulate in the plant?
Figure 3. Trees are able to draw water from the soil and transport it to the tops. [Source: royalty-free image / Pixabay]In everyday life, water circulates by gravity, that is, by the effect ofgravity. This is how we get running water from our taps, because it comes from a water tower built higher than our home. Of course, pumps are needed to fill the water tower from tanks below. These pumps are located at the reservoirs and create a pressure that pushes the water into the water tower. If you want to draw water from a well, you can either use a bucket on a chain or a pump. For a shallow well, a suction pump can be used on the surface. It creates a relative vacuum and the atmospheric pressure pushes the water from the well into this vacuum and allows it to come to the surface. But a suction pump cannot create a negative pressure, it can only draw water up to a maximum height of 10 m, corresponding to the atmospheric pressure.
Yet we see around us trees 15 to 30 m high, which seem to have no problem supplying their leaves with water (Figure 3), and there are even trees in California and Australia that are up to 100 m high. How do they do it? This has long been debated because the answer offends common sense, but it is no longer in doubt.
The water in a leaf, present in the cells and intercellular spaces, evaporates inside the leaf during the day. This loss of water causes a suction or negative pressure in the liquid water at the evaporation sites. Since water molecules are tightly bound together by electrostatic forces, this negative pressure pulls water from the soil through the roots and stem vessels to the leaves. This allows plants to suck up water from a height of more than 10 m, something that humans cannot do with a suction pump.
We have seen that water circulates by gravity and under the effect of pressure differences. The presence of dissolved salts in water also attracts water molecules by what is called osmotic force. And when water is in a porous medium such as soil or a cell wall, it is bound to the solid matrix by binding forces known as capillarity. Physicists have sought to unify these various forces in the notion of water potential or potential energy of water, with water always flowing in the direction of decreasing potential. To clarify this further, we need to go into more detail about the physical properties of water, the concept of water potential and the factors affecting transpiration.
2. Plants and water: the details
2.1. The physical properties of water
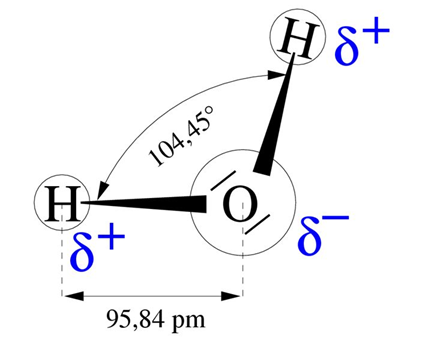
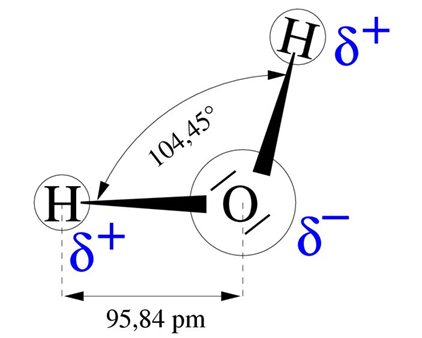
Water has remarkable properties related to its molecular structure, which is formed by two O-H bonds at an angle of 104° to each other and which form an electric dipole (Figure 4). The positive side of this dipole attracts the negative side of another molecule, ensuring the cohesion of the liquid. This cohesive force is important and explains the high energy required to evaporate water: on average 590 cal/g, or 2450 J/g or 2.45 MJ/kg. This energy, the highest of all liquids, is about 6 times the energy needed to bring water from 0 to 100°C.
Figure 4. Structure of the water molecule. 1 pm = 10-12 m. [Source: author’s diagram]The consequence is that evaporation has a strong cooling power. Thus, in summer, a dry asphalt surface has been measured to be 49°C, while the surface of a nearby pond is 26°C and the surface of a wet meadow or alder wood 29°C. Through their evaporation, plants in general and trees in particular play an important role in climate regulation. Trees have the advantage of maintaining a leaf cover throughout the growing season thanks to their deep roots, which enable them to maintain a water supply during the dry season.
Another consequence of the strong cohesion between water molecules is the high density of liquid water, which is higher than that of ice, allowing ice to float. The water also has a high surface tension, which allows gerrids (insects improperly called water spiders) to run across the surface of the ponds.
2.2. Water potential and its components
The water potential Ψ is the potential energy of water, modified by the effects of gravity, osmotic and matrix phenomena and pressure differences (See How do plants cope with a salty diet?). It is expressed in joules per cubic metre (J m-3), the dimensions of a pressure (in pascal, Pa). The pascal being a very small unit, the MPa or the bar is frequently used (1 bar = 0.1 MPa).
The gravitational potential Ψg is the gravitational potential energy m.g.h, referred to the unit volume V, where m is the mass of water, g the acceleration of gravity, and h the height from a reference level (the ground surface in general):
Ψg = (m/V).g.h = ρ.g.h
Ψg is positive if the water is above the ground (h>0), and negative otherwise (h<0). Thus in a tree, Ψg is negative for root water and positive for leaf water.
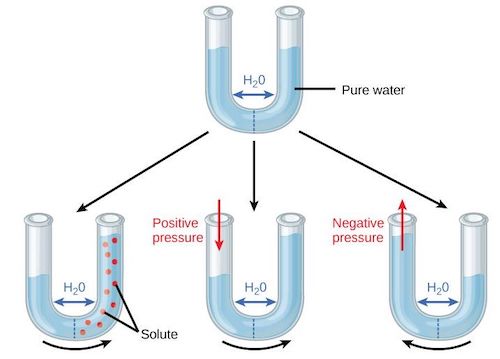
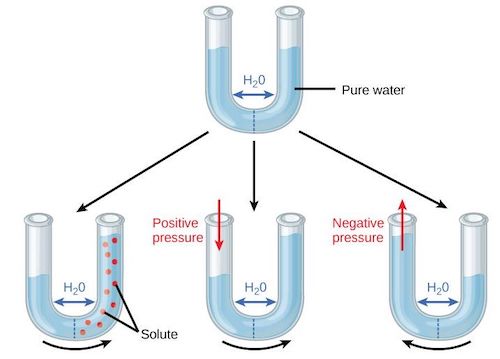
Figure 5. Top: U-tube separated by a semi-permeable wall and filled with pure water. Bottom: effects of solutes (left), pressure (center) and suction (right). [Source: author’s diagram]The osmotic potential or solute potential Ψs the potential energy of water bound to solutes. Osmotic phenomena are shown in Figure 5 using a U-tube with two halves separated by a semi-permeable wall, i.e., permeable to water but not to solutes. At the top of the figure, pure water is placed in the tube, flowing freely through the membrane and reaching an identical level between the two halves. At the bottom left, a solute (red dots) is placed in the right half, the pure water in the left half is attracted to the salt molecules and the water rises to the right half. At the bottom center, the same movement is simulated by exerting positive pressure on the water in the left half, and at the bottom right the movement is reversed if negative pressure is exerted on the water in the left half.
The osmotic potential Ψs always negative because water bound to solutes is less available than free water. It depends on the concentration of solutes Cs. To a first approximation for a dilute solution, this dependence is linear:
Ψs = – R.T. Cs
where R is the perfect gas constant and T is the absolute temperature
The matrix potential (Ψm) is the energy binding the water to the solid particles of the matrix (soil or cell wall), it is proportional to the contact surface (4πR2 for a spherical pore), and reported to the volume (4 πR3/3), it varies as 1/R, i.e. as the inverse of the pore radius. It is always negative, and all the more so as the pores are small. Thus a sandy soil with coarse pores retains little water, while a clay soil with very small pores retains much more. The soil water available to plants varies between two limits:
An upper limit: the field capacity ;
and a lower limit: the permanent wilting point.
Above the field capacity, water flows by gravity, below the permanent wilting point, residual moisture is not available to plants and corresponds to Ψm = – 15 bar (or – 1.5 MPa).
The interval between these two limits is called useful reserve, for a soil of 1 m depth, it represents between 100 and 200 mm of water height, which represents 30 to 60 days of consumption for a crop in good conditions. In practice, plants begin to close their stomata when about half of this reserve has been used up, and the reserve is lower if roots are shallower than 1 m. But trees have roots well below 1 m, thus increasing the useful reserve. It reaches 400 mm in the Barbeau sessile oak forest near Melun, according to the researchers who study it at the Laboratoire d’Ecophysiologie végétale, Université Paris-Saclay (see below).
Finally, the pressure potential Ψp is directly equal to pressure, and thus positive in a turgid cell and negative in vessels during the day.
The water potential Ψ is thus the sum of these four components, and water always flows in the direction of decreasing potentials:
Ψ = Ψg + Ψs + Ψm + Ψp
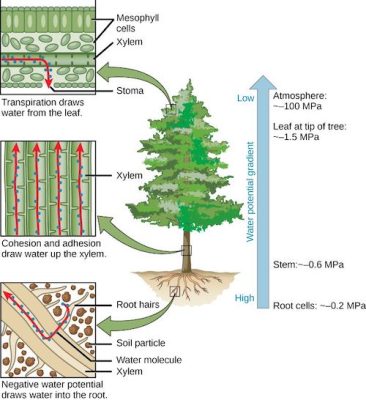
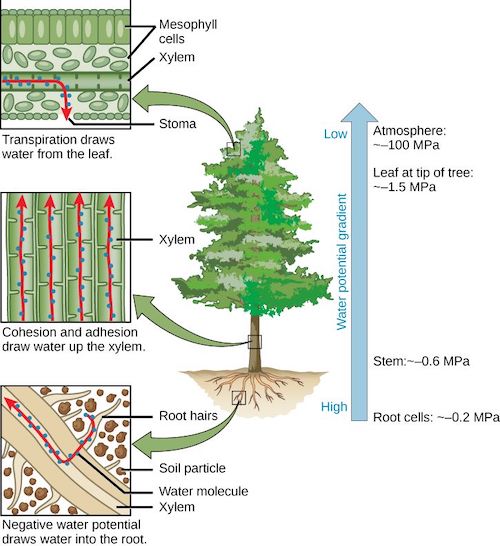
Figure 6 shows the variations of this potential in a transpiring tree during the day:
Figure 6. Water circulation in a tree according to decreasing potentials. [Source: author’s diagram]The soil potential is slightly negative (- 0.05 MPa or – 0.5 bar) because the water is below the surface and contains dissolved mineral salts in low concentrations.
At the top of the tree, the potential of a leaf is – 1.5 MPa or – 15 bar, because the leaf is subject to significant transpiration. This strongly negative value exerts suction on the entire water column, from the roots to the foliage, resulting in intermediate values for the roots (-0.2 MPa) and for the trunk (-0.6 MPa). The water column withstands these negative values of water potential (and therefore pressure) thanks to the cohesive forces that bind the water molecules together.
Note that the potential of the air (-100 MPa) is much lower than that of the sheet (-1.5 MPa). This potential is in equilibrium with the relative humidity of the air, in this case 48% in the open air, compared to 98.9% in the air inside the sheet, which is therefore almost saturated. [2]
The maximum height of trees is about 100 m, which corresponds to a gravitational potential of 10 bar or 1 MPa. This means that the leaves must be at a potential well below -1 MPa to be able to move water up to this height.
2.3. Evaporation and boundary layer, aerodynamic resistance
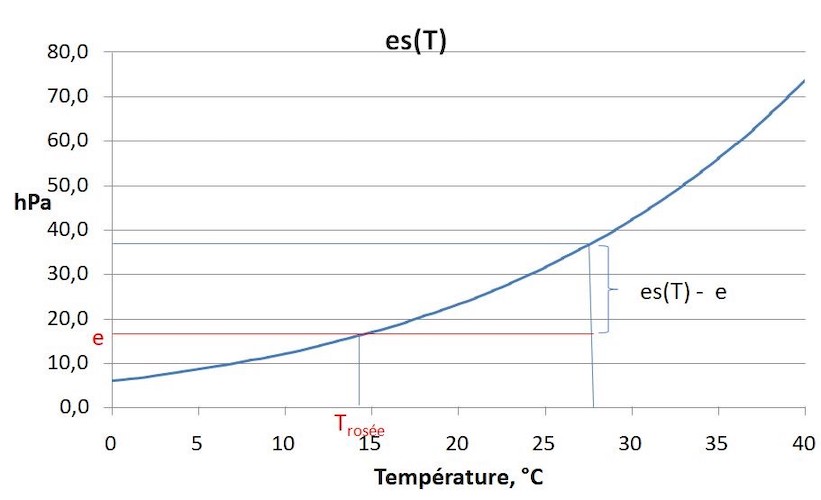
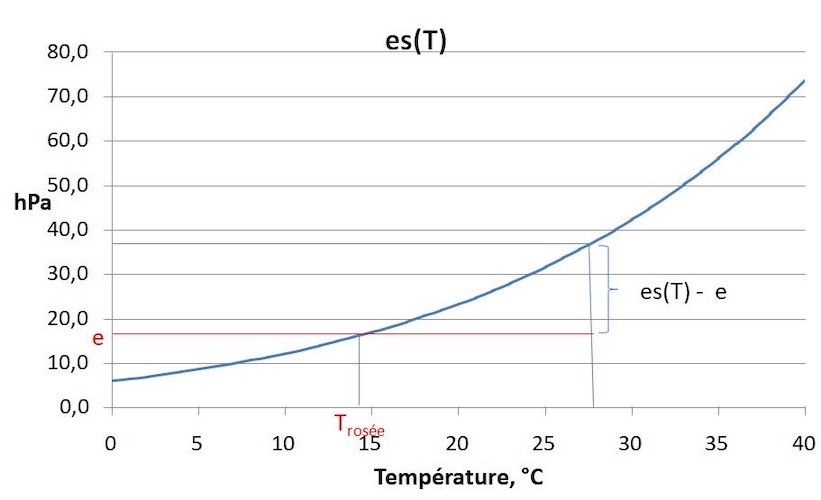
Water is present as vapour in the air. When water vapour is injected into dry air, the water vapour pressure (e) increases to a maximum value, above which the vapour condenses into liquid water. This is called the saturation vapour pressure es(T), which increases exponentially with temperature (Figure 7). The ratio e/ es (T) is the relative humidity and is expressed in %. The difference es (T) – e is the saturation deficit, which increases strongly with temperature.
Figure 7. Saturation vapor pressure of water as a function of temperature. Air with a vapour pressure e has a saturation deficit es(T) – e, and if its temperature is lowered, dew is formed at T<Td where es (Td) = e. [Source: author’s diagram]Inside a leaf, the air is practically saturated with water vapour and transpiration of a leaf is caused by the difference in vapour pressure between the inside and outside of the leaf.
Air in contact with liquid water is saturated with water vapour, and the evaporation of a free water surface E will depend on the saturation deficit [es(T) – e] between the surface and the free air. E increases with the wind speed, and we define an aerodynamicresistance ra, the resistance opposed by the air to the water vapour flow E between the evaporating surface and the free air.
Most of this resistance is located in the so-called boundary layer, which extends from the surface where the air speed is zero to the open air. Its thickness is of the order of a millimetre for a leaf, a few centimetres for a surface of open water, and can reach several decimetres for a cornfield and several metres for a forest.
2.4. Transpiration
Resistance ra can be applied to an open water surface (pond, lake) or to a wet object such as a damp blotting paper. For a plant, the water vapour must first diffuse through the epidermis and its stomata, before exiting into the outside air, which is represented by rs, a resistance that adds to ra as two resistances in series.
A leaf is often compared to a wet blotting paper that evaporates freely under the influence of physical factors. Solar radiation provides most of the evaporative energy, wind speed changes the resistance ra, air temperature and humidity affect the air saturation deficit. The higher the radiation level and the hotter and drier the air, the greater the evaporation. The difference between the leaf and the wet blotter is the presence of the epidermis and its stomata, which slows down evaporation, like a plastic sheet with small holes:
When the stomata are well open, transpiration is close to the evaporation of the blotter (but a little less);
When the stomata close, transpiration decreases to very low values due to the residual porosity of the epidermal cuticle. This results in an increase in leaf temperature which is no longer cooled by evaporation.
The flux E over a plant canopy is called evapotranspiration, which is the sum of evaporation from the soil and wetted surfaces (if any), and transpiration from leaves. It is closely related to the leaf area index, which is the ratio of the leaf area of a vegetation to the underlying soil area.
In practice, the flow of water evaporated from a natural surface depends on the surface conditions (open water, soil with varying degrees of vegetation cover and moisture) and climatic conditions (solar radiation, air temperature and humidity, wind speed).
Potential evapotranspiration Ep is the amount of water evaporated by a low vegetation surface completely covering the ground and well supplied with water. Ep varies from 0 to 10 mm/d depending on the climate; over a year Ep can go up to 2700 mm, the maximum value observed in hot and dry places like the Sahel, for well-greened irrigated grass.
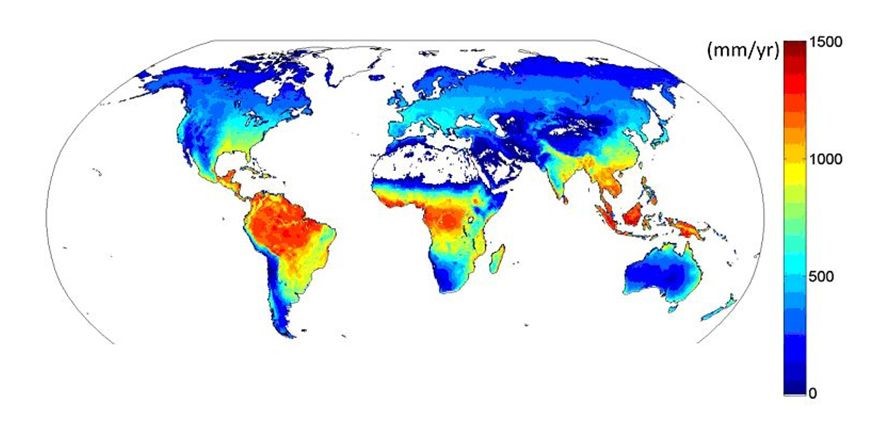
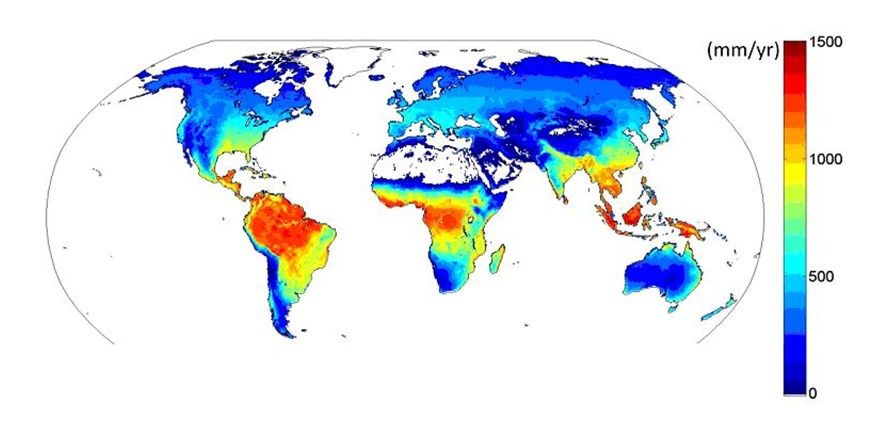
Figure 12. Simulated evapotranspiration from Fluxnet flux measurements [Source: Jung M et al. ref. [4]]Several hundred of these devices exist in the world and have allowed to establish a first map of its variations in 2010 (Figure 12) [5]. Evapotranspiration is maximal (up to 1300 mm/year) over tropical rainforests near the equator, decreases towards the dry tropics, then rises towards temperate regions before decreasing towards the poles due to low values of temperature and radiation.
3. Role of water in plant production
3.1. Is transpiration necessary for plants?
Yes, because leaves need to be open to the outside air to be able to absorb the carbon dioxide they feed on, and this is why their epidermis is pierced by numerous stomata (see What is the impact of air pollutants on vegetation?). The air inside the leaf is very humid because it is in contact with well-hydrated cells, and no plant or animal cell is known to function without water. Transpiration results from this porosity of the leaf epidermis and from the difference in humidity between the air inside and the air outside the leaf. Plants also need mineral elements other than carbon (nitrogen, phosphorus, potassium…) which they find in the soil solution absorbed by the roots through transpiration.
It is considered that to produce one kilogram of wheat, a crop must consume about 500 liter of water. This figure is an order of magnitude; it is slightly lower in a humid climate and higher in an arid climate because of variations in evapotranspiration.
3.2. Consequences for the circulation of sap
Liquid water is made up of polar molecules, which attract each other with particularly high cohesive forces. This high cohesion makes it possible for the sap to flow upwards in large trees, where the water in the vessels is under very low negative pressure. However, the water column in the vessels can break when the tension becomes too strong, and a gas bubble appears, blocking the circulation of water. This is cavitation, which is the equivalent of embolism for blood vessels. A tree has many vessels, and begins to suffer when cavitation hits a significant portion of these vessels [6]. The anatomy of vessels in relation to their sensitivity has been studied extensively.
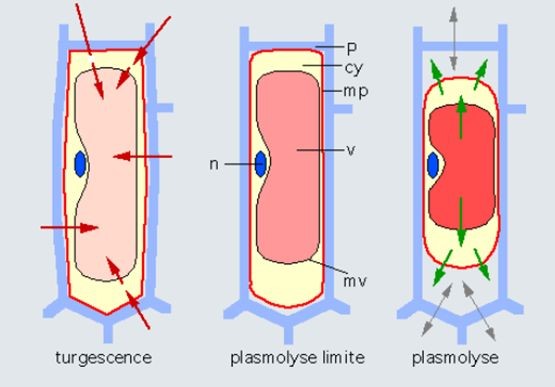
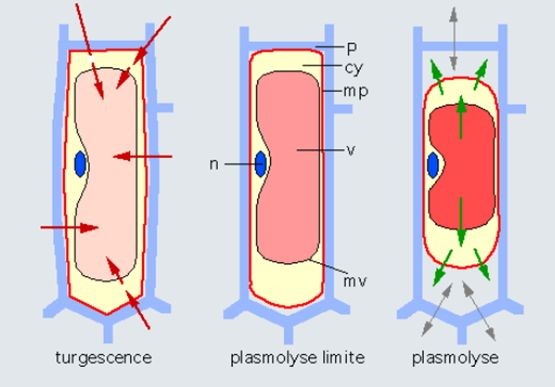
Figure 13. Effect of medium concentration on cell water status. On the left, turgid cell in a medium with low solute concentration. In the center, the medium has the same concentration as the vacuole. There is no water exchange. The cell exerts no pressure on the wall. The turgor pressure is zero. This is the limit plasmolysis. On the right, plasmolyzed cell is in a solute-concentrated medium. The arrows indicate that there is an outflow of water from the cell before a new equilibrium is reached. Abbreviations: cy, cytoplasm; mp, plasma membrane; mv, vacuolar membrane; p, wall; v, vacuole; n, nucleus. The cell part inside the wall is the protoplast. [Source: author’s diagram]Liquid water is also an excellent solvent, surrounding each mineral ion with a shell of water molecules. The forces that connect solute molecules to water molecules are called osmotic forces, and the more concentrated the solution, the more water it attracts, causing a buoyancy (osmos means buoyancy in Greek).
The water in a plant is found in the cells that make up the plant, and also in the vessels that feed the plant from the roots to the leaves. The water in the soil is generally low in mineral elements, whereas the water in the cells is quite concentrated. This difference in concentration attracts a flow of water from the soil to the plant by osmosis, which swells the cells (Figure 13, left). The water is therefore normally under pressure in the cells, and the cells are said to be turgid. This pressure is contained by the cell walls, which are made up of relatively rigid cellulose and pectin fibres, which act like a tyre that contains the pressure of an inner tube. A car tyre is inflated to between 2 and 3 bars, a bicycle tyre can be inflated to 6 bars, and the pressure can be as high as 10 bars in a plant cell. It is because of these high pressures that plant cells have a wall, which does not exist in animal cells, which are subject to lower pressures. This pressure decreases when the cell dries out, and then is cancelled out, as the cell membrane peels away from the wall, which is known as plasmolysis (Figure 13, centre and right).
4. Messages to remember
Water is the necessary medium for all forms of life
It is under pressure in plant cells, and this pressure allows growth and ensures the shape and rigidity of plants, as well as the lignocellulosic tissues of stems and trunks.
On the other hand, it is under tension in the leaves and vessels, and this tension allows the rise of the sap, which compensates for the loss of water from the leaves through transpiration and evaporation.
Transpiration takes place mainly through the stomata, the pores necessary to ensure the absorption of carbon dioxide.
Stomata are wide open in wet soil and close in dry soil to conserve internal water, thus inhibiting CO2entry and thus photosynthesis and growth.
Stomatal closure is triggered by the drop in water potential which, when it becomes strong, also causes the formation of gas bubbles in the vessels by cavitation.
The mechanisms of plant adaptation to water shortage are discussed in the associated article (forthcoming).
Notes and references
Cover image. Vineyard in the park of La Geria on the island of Lanzarote (Canary Islands). The rare rains (170 mm/year) are retained by the low stone walls and give a quality wine. [Source: Patrice78500, CC BY-SA 4.0, via Wikimedia Commons]
[2] The formula relating the potential to atmospheric humidity is: Ψ = (RT/Vwater)*ln(e/es(T)) where R is the perfect gas constant, T is the absolute temperature, Veau is the volume of one mole of liquid water and e/es(T) is the relative humidity. Taking R = 8.3 J mol-1 K-1, T = 298 K, Veau = 18.10-6 m3 mol-1 and e/es(T) = 0.48 gives Ψ = – 100 MPa. Thus the water potential of air is generally very low, except when the air is almost saturated (hr>98%) as in the air inside the leaf.
3] Dufrêne E, Pontailler J-Y and Saugier B, 1993. A branch bag technique for simultaneous CO2 enrichment and assimilation measurements on beech (Fagus sylvatica L.) Plant, Cell and Environment, 16, 1131-1138
[5] Jung M et al. , 2010. Recent decline in the global land evapotranspiration trend due to limited moisture supply. Nature467, 951-954.
[6] The interested reader will find more information in the excellent presentation (only in french) of André Granier and Hervé Cochard in the website “Les plantes et l’eau : II.7. L’architecture hydraulique, la cavitation et l’embolie des plantes”.