


Symbiosis and evolution: at the origin of the eukaryotic cell
PDF
Over two billion years ago, life was exclusively microbial, inhabiting oceans, marine sediments and hydrothermal environments. In these environments, close associations developed between microorganisms, some of which evolved into true endosymbiotic relationships, involving the long-term integration of one organism within another. Among these events, one would profoundly transform the history of life: the association between an archaeon related to modern Asgard archaea and an α-proteobacterium, which gave rise to mitochondria and the eukaryotic lineage. From this alliance emerged animals, plants, fungi and a vast diversity of single-celled eukaryotes. The eukaryotic cell is characterised by its complex organisation, with a nucleus containing most of the DNA, mitochondria responsible for much of the cell’s energy production and, in plants and many algae, chloroplasts that carry out photosynthesis. Subsequent endosymbiotic events further shaped evolution: the acquisition of a cyanobacterium gave rise to primary plastids, including chloroplasts, whilst secondary and tertiary endosymbiosis events spread plastids among numerous lineages. Endosymbiosis thus emerges as a major driving force behind evolutionary innovation and the diversification of life.
1. The eukaryotic cell is a chimera
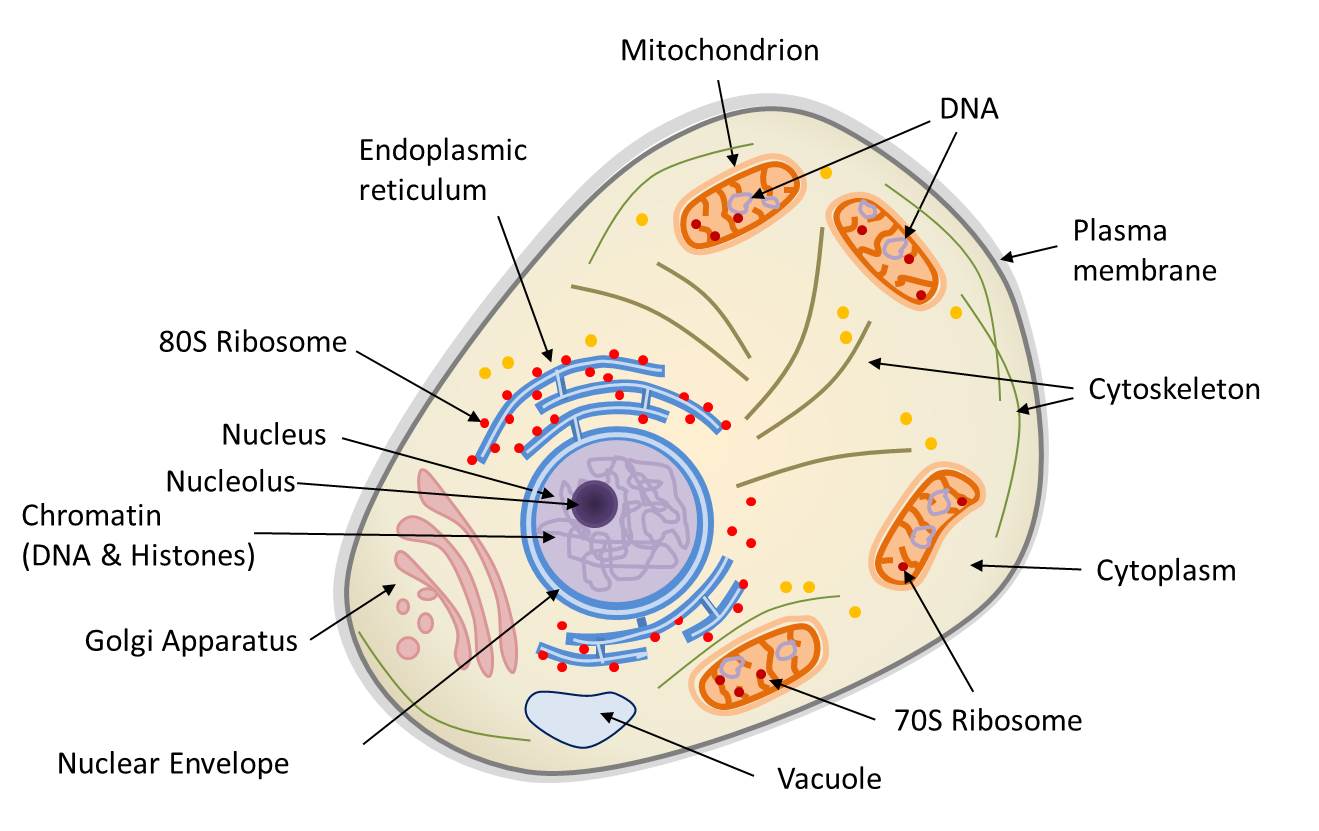
The eukaryotic nucleus is bounded by a double membrane, the nuclear envelope (Figure 1). It contains the nuclear genome, which consists of DNA and carries the vast majority of the genes required for cellular function, carried by DNA (deoxyribonucleic acid). However, the eukaryotic cell also harbours non-nuclear genomes within its organelles:
- the mitochondrial genome, located in the matrix of the mitochondria (Figure 1);
- the chloroplast genome, located in the stroma of the chloroplasts (in plants and algae).
The DNA molecules that constitute these three genomes are not organised in the same way. In the nucleus, the genome is distributed across several linear DNA molecules – the chromosomes – which are associated with proteins called histonesBasic proteins that combine with DNA to form the basic structure of chromatin. Histones play an important role in DNA packaging and folding.. It contains both coding sequences (transcribed into messenger RNA and then translated into proteins) and non-coding sequences. The three-dimensional configuration of the nuclear genome is functionally crucial: the folding of DNA and its wrapping around histonesBasic proteins that combine with DNA to form the basic structure of chromatin. Histones play an important role in DNA packaging and folding. allows a large amount of genetic information to be packed into a very small volume. In contrast, mitochondrial and chloroplast genomes are generally composed of circular DNA molecules, organised differently from nuclear DNA. In all cases, DNA retains its double-helix structure; only the higher-level organisation and overall topology differ: organellar DNA is generally circular, whereas nuclear chromosomes are linear.
Indeed, prokaryoticProkayotes are microorganisms (usually unicellular) with a simple cellular structure, no nucleus, and almost never internal compartmentalization (the only exception being thylakoids in cyanobacteria). Currently, living organisms are considered to consist of three groups: Archaea, Bacteria (Prokaryotes) and Eukaryotes. cells (bacteria and archaeaUnicellular microorganisms devoid of nucleus and living in extreme environments (anaerobic, high salinity, very hot…). Phylogenetic research by Carl Woese and George E. Fox (1977) differentiated between archaea and other organisms. Currently, living organisms are considered to consist of three groups: archaea, bacteria and eukaryotes.) do not possess a true nucleus. Their DNA is most often circular, as are most mitochondrial and plastid genomes. This organisation suggests that these organelles may have evolved from ancient prokaryotes. Transcription and translation therefore occur without separation by a nuclear envelope; DNA replication also takes place outside a nucleus. It should be emphasised, however, that archaea are only superficially comparable to bacteria: many aspects of their cell biology and metabolism differ profoundly, and the mechanisms of replication, transcription and translation involve proteins and processes more closely resembling those of eukaryotes. Finally, prokaryotes generally exhibit limited and less complex internal compartmentalisation (cyanobacteria being a notable exception). Above all, their compartments, where they exist, are not mobile: their cytoskeletal systems, where present, are generally less elaborate and do not support organelle trafficking comparable to that of eukaryotes.
Table 1. Comparison of eukaryotic and prokaryotic cells.

Table 1 compares the properties of prokaryotic and eukaryotic cells, taking into account the mitochondria and, where applicable, the chloroplasts of the latter. It shows that mitochondria and chloroplasts share many features with prokaryotic cells. Apart from the structure of their DNA, the organelles of eukaryotic cells arise only from pre-existing organelles and multiply by binary fission, just like bacteria. They also retain bacterial-type protein-synthesis machinery: 70S-type ribosomesHuge complex composed of RNA and ribosomal proteins, associated with a membrane (at the granular endoplasmic reticulum) or free in the cytoplasm. The function of the ribosome is to translate messenger RNA (mRNA) into proteins. The enzymatic activity of the ribosome being carried by rRNAs, the ribosome is a ribozyme. Common to all cells (prokaryotes and eukaryotes), the structure and composition of the ribosome varies according to the organisms. In prokaryotes and cellular organelles (mitochondria, chloroplast), the ribosome is said to be 70S (S corresponding to the so-called Svedberg sedimentation unit) and consists of the 50S and 30S subunits. The ribosome of eukaryotes is called 80S and consists of two subunits 60S and 40S., which occur free in the matrix or stroma, whereas the eukaryotic cytoplasm utilises 80S-type ribosomes, sometimes attached to the membranes of the endoplasmic reticulumMembrane network of the cytoplasm of eukaryotic cells, essential for cellular metabolism (lipid and protein synthesis, calcium storage). Associated with ribosomes, it is the place of synthesis of proteins secreted outside the cell and, on the other hand, proteins and lipids constituting the membranes of cellular organelles (Golgi apparatus, lysosomes, mitochondria, nucleus, ribosomes, vesicles…).. Finally, certain bacteria carry out metabolic processes related to those of mitochondria (respiration) and chloroplasts (photosynthesis). The eukaryotic cell, on the other hand, is distinguished by the cytoskeleton, a self-organising and mobile protein system that positions and moves organelles within the cell. In prokaryotes, such a network is much simpler and far less dynamic than that of eukaryotes and remains poorly developed in mitochondria and chloroplasts.
These properties show that the eukaryotic cell is a biological chimera, that is to say, an organism whose genetic heritage derives from several distinct evolutionary lineages: it combines components of archaeal origin (notably much of the informational machinery associated with the nucleus) with components of bacterial origin (chloroplasts, mitochondria).
As early as the turn of the twentieth century, the symbiotic origin of eukaryotic cells had already been proposed, but it was not until the advent of new methods for studying the cell — electron microscopy, biochemistry, molecular biology — that the endosymbiotic theory was revived, around 1970, by the American microbiologist Lynn Margulis [2] (see Focus The pioneers of the endosymbiotic theory).
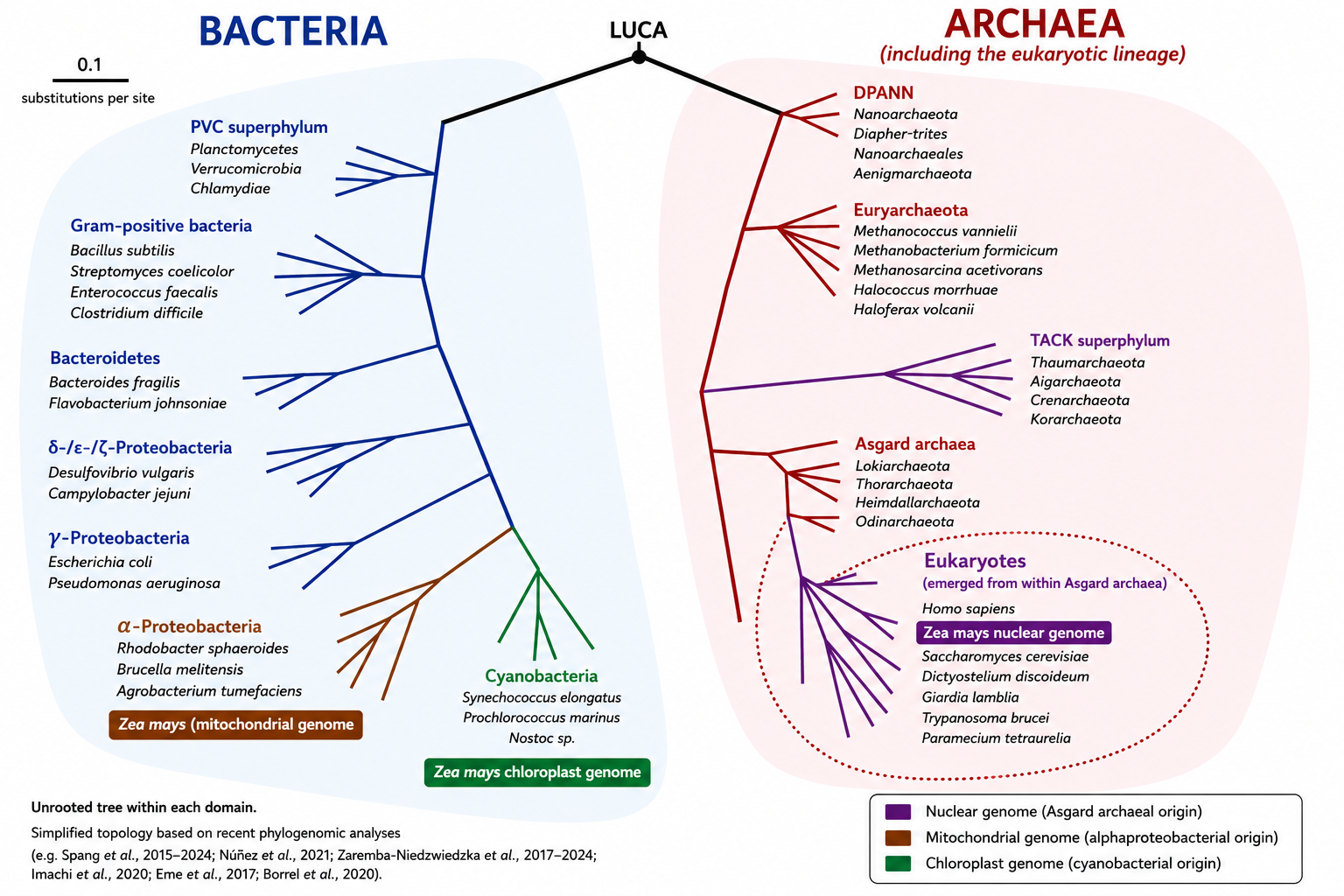
But the discovery that eukaryotes evolved from a specific lineage of archaea (Asgard) has overturned the traditional division of life into prokaryotes and eukaryotes (see Focus The Asgard Archaea Revolution).
2. How did the eukaryotic cell arise?
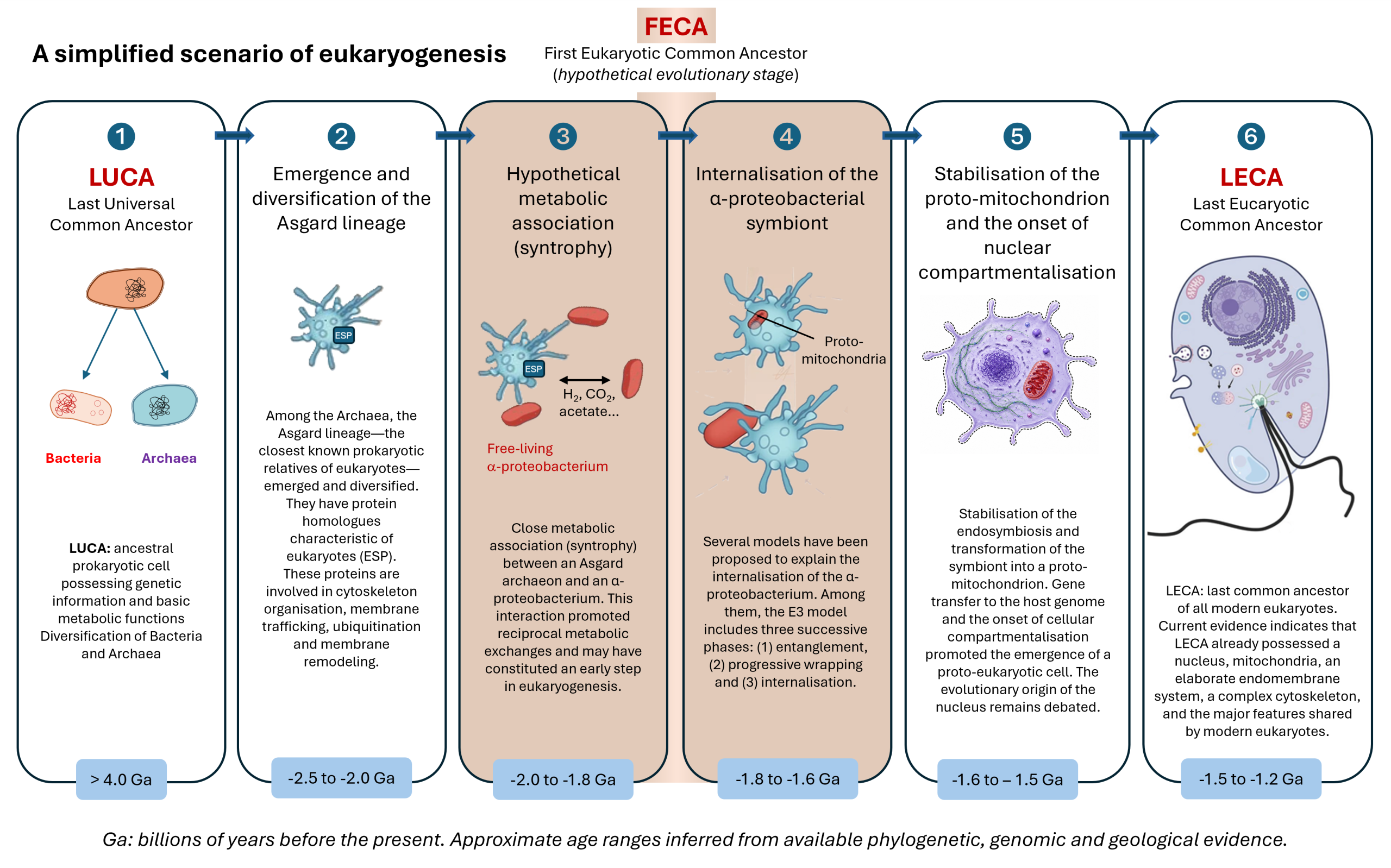
The emergence of the first eukaryotic cells, around 1.5-2 billion years ago, represents one of the most significant evolutionary transitions in the history of life. This transition occurred approximately 1.5–2.0 billion years after (see Focus LUCA, LECA and the common ancestors of the tree of life), it enabled the emergence of unprecedented cellular complexity and paved the way for the evolution of multicellularity. Data from genomics, phylogenomics and microbiology now converge on this scenario, which is currently the best-supported framework for explaining the origin of eukaryotic cells.
2.1. The host cell: an Asgard archaeon


Recent reconstructions of the last common eukaryotic ancestor (LECA; see Focus LUCA, LECA and the common ancestors of the tree of life) show that the vast majority of conserved genes of non-mitochondrial origin originated from an archaeal lineage related to modern Asgard archaea, with a dominant contribution in most cellular functional systems (replication, transcription, translation, cytoskeleton, endomembranes). LECA was already a complex cell, possessing a nucleus, a dynamic cytoskeleton, a well-developed endomembrane system, phagocytic capabilities and, probably, mechanisms of sexual reproduction (meiosis) [4],[5]. Figure 4 schematically summarises these cellular characteristics of LECA [6].
2.2. The symbiotic scenario: metabolic and molecular mechanisms of H₂ syntrophy
One of the most widely discussed scenarios (Figure 5) [7] is based on a metabolic symbiosis (syntrophyMetabolic cooperation between microorganisms based on the exchange of compounds such as H₂, CO₂, formate or acetate, the consumption of which by one partner enables the metabolism of the other. This type of interaction is regarded as a key step in theories concerning the origin of the eukaryotic cell.) between an H₂-dependent Asgard archaeon and a facultatively anaerobic α-proteobacterium [8]. This scenario is now supported by several lines of evidence, but it is not the only one proposed. Other models differ regarding the mechanisms that led to the integration of the bacterium into the host cell, whilst recognising the central role of mitochondrial endosymbiosis.
Mechanisms of H₂ syntrophy
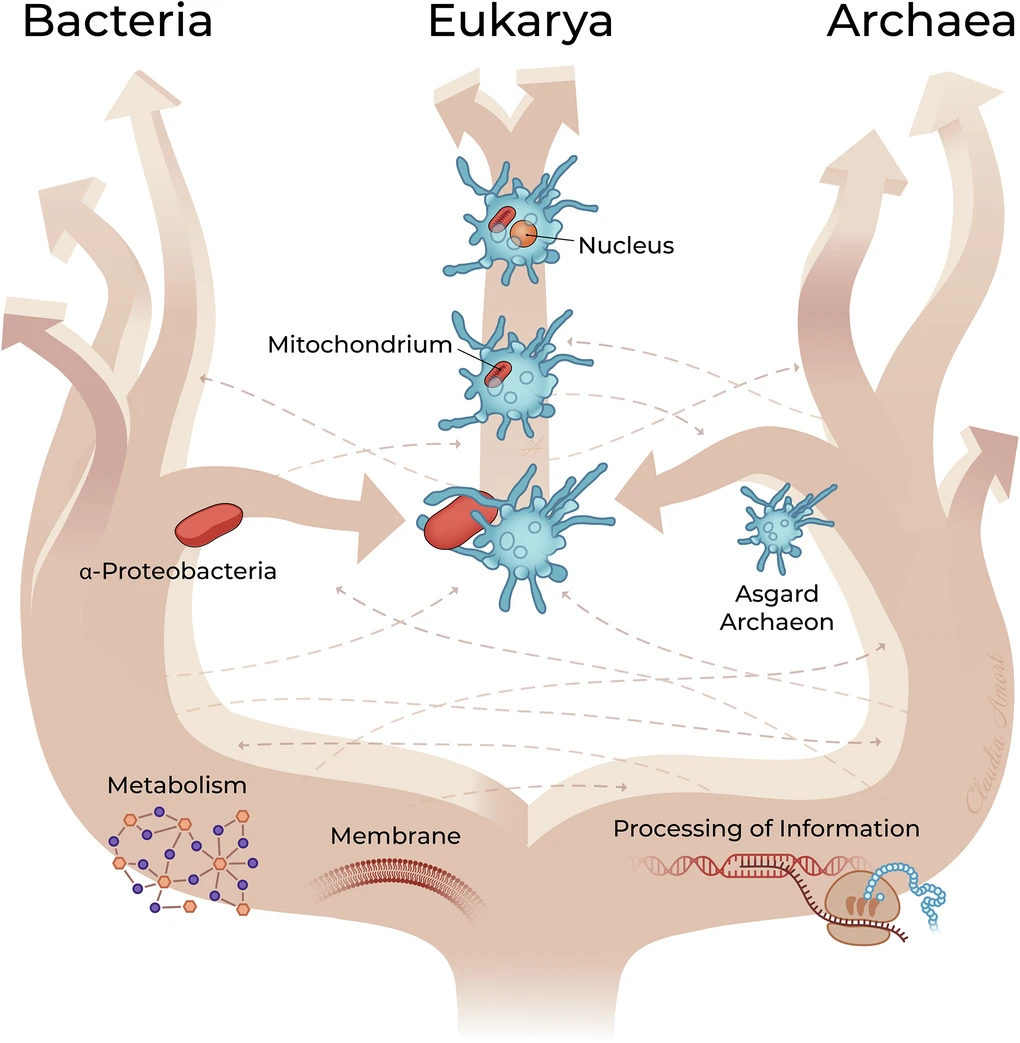
In Asgard archaea, this syntrophic metabolism involves [FeFe] or [NiFe] hydrogenases. The two partners could also exchange electrons directly through close membrane contacts, reinforcing their metabolic cooperation. The host archaeon may have been an acetogen or methanogen, may have depended strongly on externally supplied H₂. The ancestral α-proteobacterium was capable of H₂-producing fermentation under anaerobic conditions and respiration in the presence of oxygen. This flexibility enabled an increasingly close association, progressing from extracellular syntrophy to endosymbiosis [8].
Internalisation may have followed the Entangle–Engulf–Endogenize (E³) model, proposed on the basis of observations made in Candidatus Prometheoarchaeum syntrophicum (see Figure 5) [11]. According to this model, the membrane protrusions of the archaeon may have gradually enveloped the bacterial partner before internalisation by a still-unknown process, distinct from classical phagocytosis (archaea lack any known phagocytic machinery).
The integration of the α-proteobacterium profoundly transformed the host cell. By becoming the mitochondrion, it enabled the host to generate far more energy than fermentation alone, a major energy-yielding pathway in early anaerobic organisms. Mitochondria ca produce roughly an order of magnitude more ATP – the cell’s primary energy molecule – per molecule of glucose. This increase in energy facilitated the growth of cells and the emergence of complex structures, such as the cytoskeleton, the endomembrane system and other features of eukaryotic cellular complexity.
Unlike plastids, which spread through several primary, secondary and tertiary endosymbiotic events, mitochondria appear to have emerged only once in the history of life: this single acquisition constitutes the founding event from which all extant eukaryotic lineages descend.
2.3. Gene transfers: a massive and continuous process
This integration was accompanied by extensive gene transfer from the bacterium to the host’s nuclear genome (see Figure 5). The vast majority (more than 99% in many estimates) of the genes of the α-proteobacterial ancestor were either lost or transferred to the nucleus [12]:
- relocation of bacterial DNA fragments into the nucleus;
- acquisition of eukaryotic regulatory sequences and a mitochondrial targeting sequence;
- loss of the redundant copy of the organelle gene.
These stages do not occur simultaneously but over the course of a long evolutionary process. The transferred genes included metabolic genes and genes whose products support mitochondrial replication. They have considerably enriched the nuclear genome, helping to turn it into a veritable genetic chimera [13], whilst making the organelle dependent on the nucleus for the majority of its proteins. This process has continued over hundreds of millions of years and constitutes a major example of evolution by endosymbiosis.
2.4. The debate on oxygen and chronology
The exact role of oxygen in this evolutionary transition remains a matter of debate. The debate centres less on the existence of the symbiosis than on the environmental conditions under which it occurred and on the metabolism of the two partners at the time of their association. Phylogenetic reconstructions point to a common anaerobic, H₂-dependent ancestor [9]. The ancestral α-proteobacterium was itself a facultative anaerobe. However, recent studies indicate that some Asgard archaea can tolerate low concentrations of oxygen. The mitochondrion would therefore not have been merely a response to the global increase in atmospheric oxygen, but rather an energy amplifier that enabled oxygen to be used efficiently where it was locally available.
Some elements of eukaryotic cellular complexity may have emerged before the acquisition of mitochondria in the Asgard host, as suggested by the presence of ESP-homologous proteins and certain cellular characteristics of modern Asgard archaea. The debate over the precise chronology remains open.
2.5. What about the nucleus?
The question of the origin of the eukaryotic cell is also linked to that of the nucleus. It is one of the most intensely debated questions in eukaryogenesis. The autogenous hypothesis proposes that the nuclear envelope arose through the gradual invagination of the plasma membrane of the archaeal host (without the intervention of an external partner), whereas the symbiogenic hypotheses, including some viruses, propose that it derives from the fusion or internalisation of another organism — a bacterium, or in some versions a DNA virus — whose membranes or cellular organisation may have contributed to nuclear compartmentalisation. This separation between the nucleus and the cytoplasm may have constituted an evolutionary response to the need to separate RNA splicing from translation, a process that became particularly important following the massive transfer of genes into the nucleus. Selection may then have favoured to the establishment of this compartmentalisation [9].
The eukaryotic cell is therefore the result of a profound chimerisation: an Asgard-related archaeal host contributing much of the informational machinery and part of the cytoskeleton, an α-proteobacterium contributing oxidative metabolism via the mitochondrion following an initial H₂-based syntrophicSyntrophy is a metabolic cooperation between microorganisms based on the exchange of compounds such as H₂, CO₂, formate or acetate, the consumption of which by one partner enables the metabolism of the other. This type of interaction is regarded as a key step in theories concerning the origin of the eukaryotic cell.association, and massive gene transfers fusing the two lineages [8], [9] (Figure 6). This association may have relaxed energetic constraints on cellular complexity and enabled the emergence of unprecedented cellular complexity.
Unlike the origin of mitochondria, which is widely accepted, the origin of the nucleus remains one of the major unresolved questions in evolutionary biology. Several hypotheses are currently being investigated, though none has yet gained consensus.
3. The endosymbiotic origin of the chloroplast
3.1. From phagocytosis to endosymbiosis
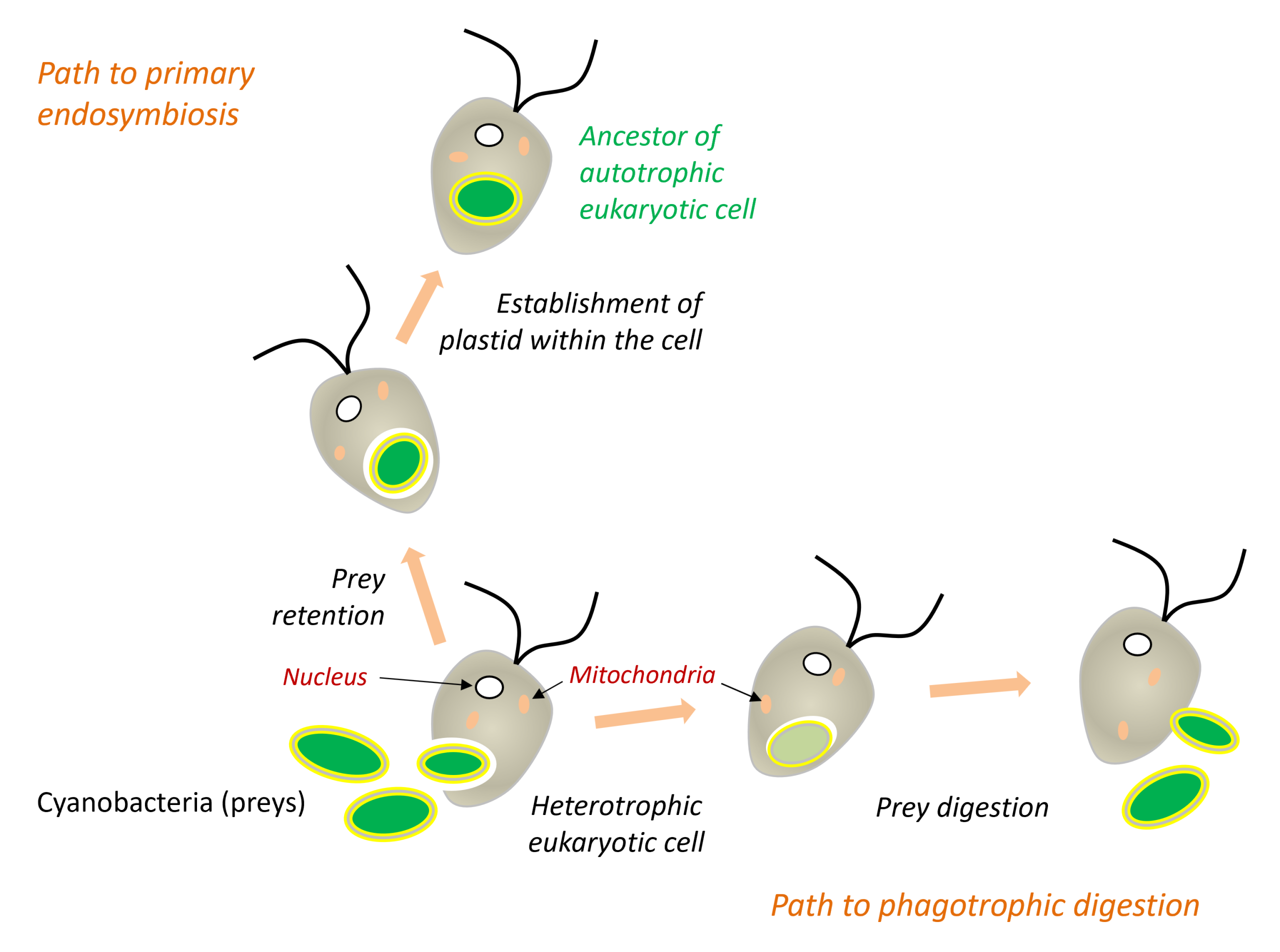
During phagocytosis, the plasma membrane invaginates around the prey and forms an endocytic vesicle, which then fuses with the lysosomes where its contents are digested. It was long thought that the outer membrane of chloroplasts originated from this endocytic membrane. Current evidence, however, suggests a more complex scenario (Figure 8). The bacterial ancestors of chloroplasts (cyanobacteria) and mitochondria (α-proteobacteria) were already GramBacteria highlighted by a staining technique called Gram staining, they appear pink under the microscope. The staining technique is based on the membrane and wall characteristics of the bacteria. However, it is not a phylogenetic classification factor as the groups “Gram +” and “Gram -” are both non-monophyletic.-negative bacteria possessing two membranes. Furthermore, the outer membrane of chloroplasts, particularly the side facing the cytosol, contains glycolipids characteristic of cyanobacteria[16]. These observations suggest that the endocytic membrane may have disappeared during the integration of the endosymbiont (Figure 8).
This remarkable longevity was long attributed to a transfer of genes from the algae into the slug’s genome. However, more comprehensive genome analyses of Elysia chlorotica have revealed no such transfer. The persistence of these ‘stolen’ chloroplasts therefore remains largely unexplained [17]. This example illustrates how a scientific hypothesis can be revised in the light of new data.
3.2. Primary, secondary and tertiary endosymbiosis
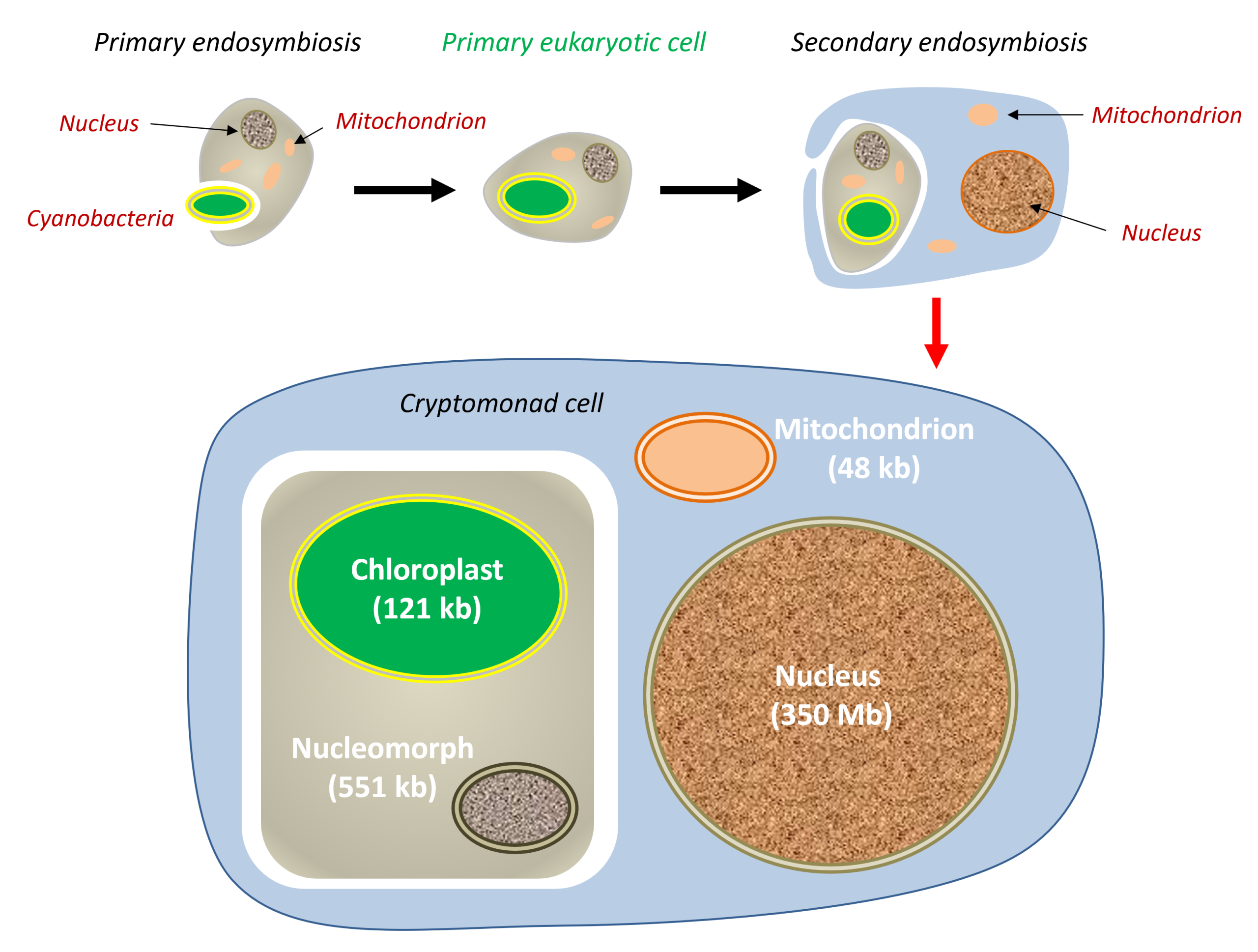
Endosymbiosis has occurred repeatedly. Independent events contributed to the origin and diversification of several eukaryotic lineages. A primary endosymbiosis involves the internalisation of a living prokaryote by a eukaryotic cell. The chloroplasts of glaucophytes, red algae and the green lineage all derive from such an endosymbiosis involving a cyanobacterium. In certain eukaryotes adapted to anaerobic environments, mitochondria have evolved significantly without ever disappearing. They have given rise to derived organelles, such as hydrogenosomesAn organelle derived from the mitochondrion, found in certain eukaryotes adapted to oxygen-free environments. It produces energy in the form of ATP through anaerobic metabolism, releasing hydrogen (H₂), illustrating the evolutionary diversity of organelles arising from mitochondrial endosymbiosis. It is found in certain anaerobic ciliates, Trichomonas, and fungi. that generate ATP anaerobically and may produce H₂ (for example, in certain ciliates) [18], as well as highly reduced organelles that retain essential biosynthetic functions, known as mitosomes [18]. Today, these organelles are grouped under the term MROA group of organelles derived from the evolution of mitochondria in eukaryotes adapted to oxygen-poor environments (anaerobic or microaerobic conditions). MROs demonstrate the remarkable evolutionary plasticity of mitochondria, which have retained or modified their functions in response to environmental constraints. Depending on the species, they perform various metabolic functions, such as ATP production (hydrogenosomes) or the biosynthesis of iron-sulphur centres (mitosomes). (mitochondrion-related organelles), which encompasses a continuum of forms derived from mitochondria. Some MROs, such as those found in Blastocystis, exhibit intermediate characteristics [19].
Secondary endosymbiosis refers to a further endosymbiotic event, whereby a eukaryotic cell that already contains an endosymbiont is internalised by another eukaryotic cell (Figure 8). This is the origin of plastids surrounded by more than two membranes, found in several eukaryotic lineages: the acquisition of a green alga by the ancestor of euglenids; the acquisition of a red alga by the ancestor of the stramenopile lineage that includes brown algae. Tertiary endosymbiosis, which is less common, has also been described. These events gave rise to new evolutionary lineages [20],[21].
Paulinella chromatophoraPaulinella chromatophora A freshwater amoeba that possesses a photosynthetic organelle derived from a cyanobacterium acquired through primary endosymbiosis around 100 million years ago. It is a unique example of primary endosymbiosis that is independent of the process that gave rise to the chloroplasts in plants and algae. a second primary endosymbiosis. This independent event, entirely independent of the one that gave rise to the plastids of glaucophytes, red algae and the green lineage, has been discovered in the amoeba Paulinella chromatophora. This amoeba descends from an ancestor that internalised a cyanobacterium related to the genera Prochlorococcus and Synechococcus, which has become a photosynthetic organelle known as a chromatophore.
Much more recent (approximately 60–140 million years ago) than the endosymbiosis that gave rise to conventional chloroplasts (over 1.5 billion years), this association is now a preferred model for studying the early stages of the transformation of an endosymbiont into an organelle[22] .
3.3. From endosymbiont to organelle
Whichever scenario is considered, the success of endosymbiosis subsequently depends on the gradual integration of the endosymbiont into the functioning of the host cell. This progressive integration transforms the endosymbiont into a true organelle. The process affects the endosymbiont’s genome, functions and transmission within the host cell.
4. The integration of endosymbionts into eukaryotic cells
4.1. A massive reduction in genome size
All the endosymbiosis events described above share a common feature: a significant reduction in the endosymbiont’s genome. Compared with the genomes of free-living proteobacteria, mitochondria have lost more than 99 per cent of their genes. Plastid genomes in the green lineage generally retain only around a hundred genes, compared with several thousand in free-living cyanobacteria. At the extreme end of the spectrum, many hydrogenosomes and all known mitosomes have lost their genomes, whilst some highly reduced plastids, such as those of the parasitic plant Rafflesia, no longer possess one either [23].
However, this genome reduction does not reflect a simplification in the functioning of the organelles. On the contrary, it reflects increasing functional integration with the host cell, with many functions now being carried out by nuclear genes.
This reduction results in particular from the loss of genes that have become redundant in the intracellular environment. Thus, cyanobacteria, like all Gram-negative bacteria, possess a layer of peptidoglycan situated between their two membranes. Although essential in the external environment, this structure becomes redundant once the endosymbiont is integrated into the host cell’s cytoplasm, where osmotic conditions are more stable. Genes required for peptidoglycan synthesis have therefore been lost from most plastids, although glaucophyte plastids retain a conspicuous peptidoglycan layer [24].
4.2. The transfer of functions and genes to the nucleus
Despite the significant reduction in their genome size, organelles retain complex proteomes, many components of which are homologous to bacterial proteins. This apparent contradiction can be explained by the fact that many proteins are now encoded by the host cell’s nuclear genome. Synthesised in the cytosol, they are then imported into the organelle via a targeting sequence, often termed a transit peptide. This relocation of functions to the nucleus constitutes one of the major innovations of endosymbiotic integration.
The systems responsible for this import represent a remarkable evolutionary innovation in their own right. Composed of proteins of both bacterial and eukaryotic origin, they recognise the proteins destined for the organelle, transport them across organelle membranes and direct them to their functional compartment [25].
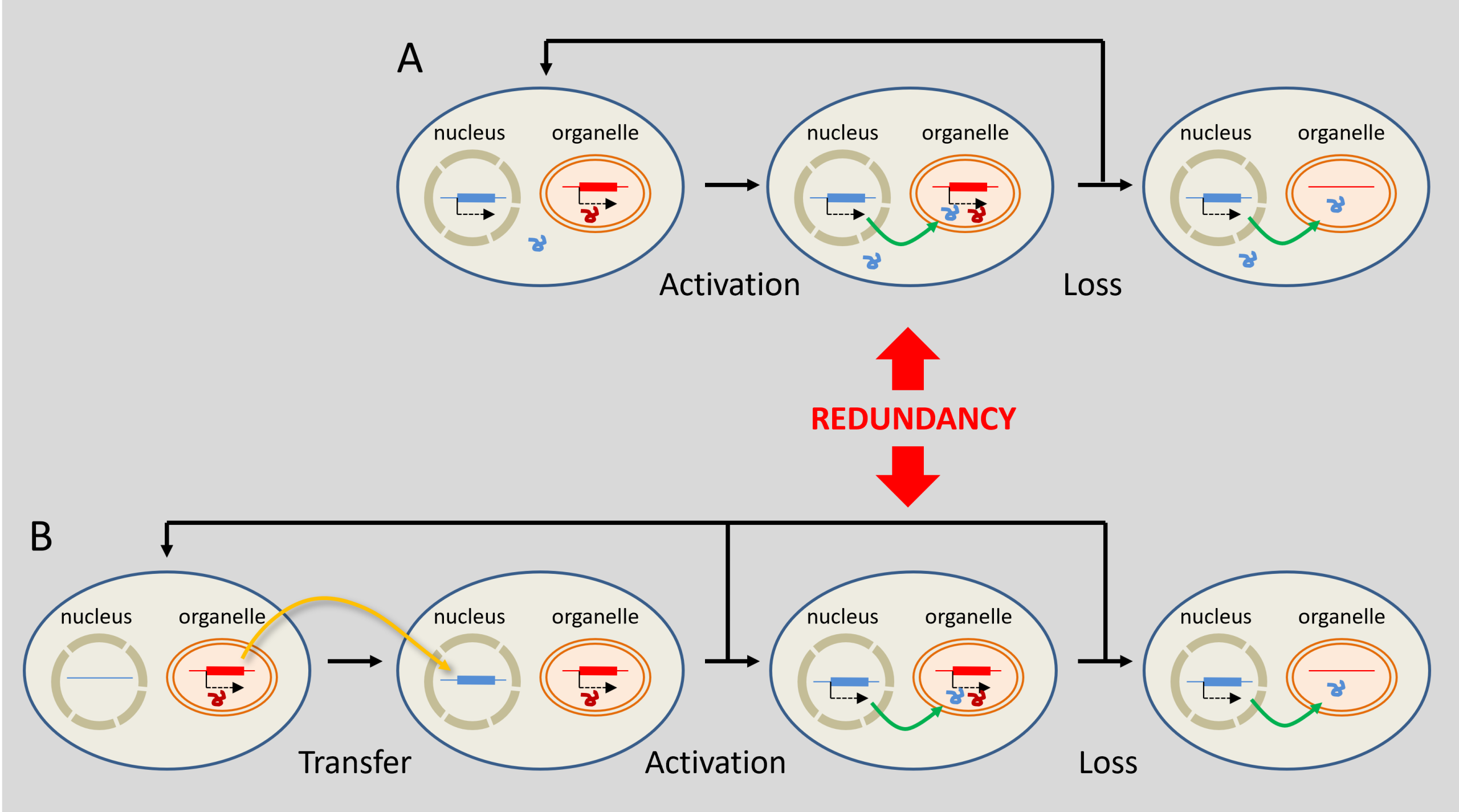
DNA transfers from organelles to the nucleus are by no means exceptional. In Arabidopsis thaliana, it is estimated that nearly 10% of nuclear genes may derive, directly or indirectly, from ancient transfers originating in plastids, often followed by duplications [26]. Their integration is thought to result from the degradation of organelles, releasing DNA fragments that are subsequently integrated into the nuclear genome.
The reduction in the genome size of organelles is the result of a balance between several evolutionary forces. Many genes have been transferred to the nucleus, whilst others have been retained in the organelles due to functional constraints that are still the subject of debate. Thus, endosymbiosis gradually reduces the genomes of the endosymbionts whilst increasing the genetic contribution to the nuclear genome, which becomes a true chimeric genome combining genes from different evolutionary lineages.
4.3. Coordination between the organelle and the host cell
The integration of an endosymbiont does not depend solely on gene transfers. It also requires close coordination between the division of the organelle and that of the host cell, an essential condition for its transmission across generations.
To be inherited by daughter cells, plastid division must be coordinated with host-cell division and be distributed between the resulting daughter cells. If they multiplied too rapidly, they would proliferate at the host’s expense; conversely, insufficient division would lead to their disappearance. Evolution has therefore favoured close coordination between the division cycles of the host and the endosymbiont.
Most of the proteins involved in chloroplast division are derived from the division machinery of cyanobacteria. A few, however, are of eukaryotic origin and, remarkably, the components discussed here are now encoded by the nuclear genome. This nuclear control constitutes one of the clearest manifestations of the endosymbiont’s integration into the host cell.
The integration of an endosymbiont is therefore not limited to its retention within the cell or to the transfer of some of its genes. It results in a genuine functional fusion between two previously independent partners. This innovation has profoundly transformed the evolution of eukaryotes.
5. Symbiosis: a driving force in evolution?
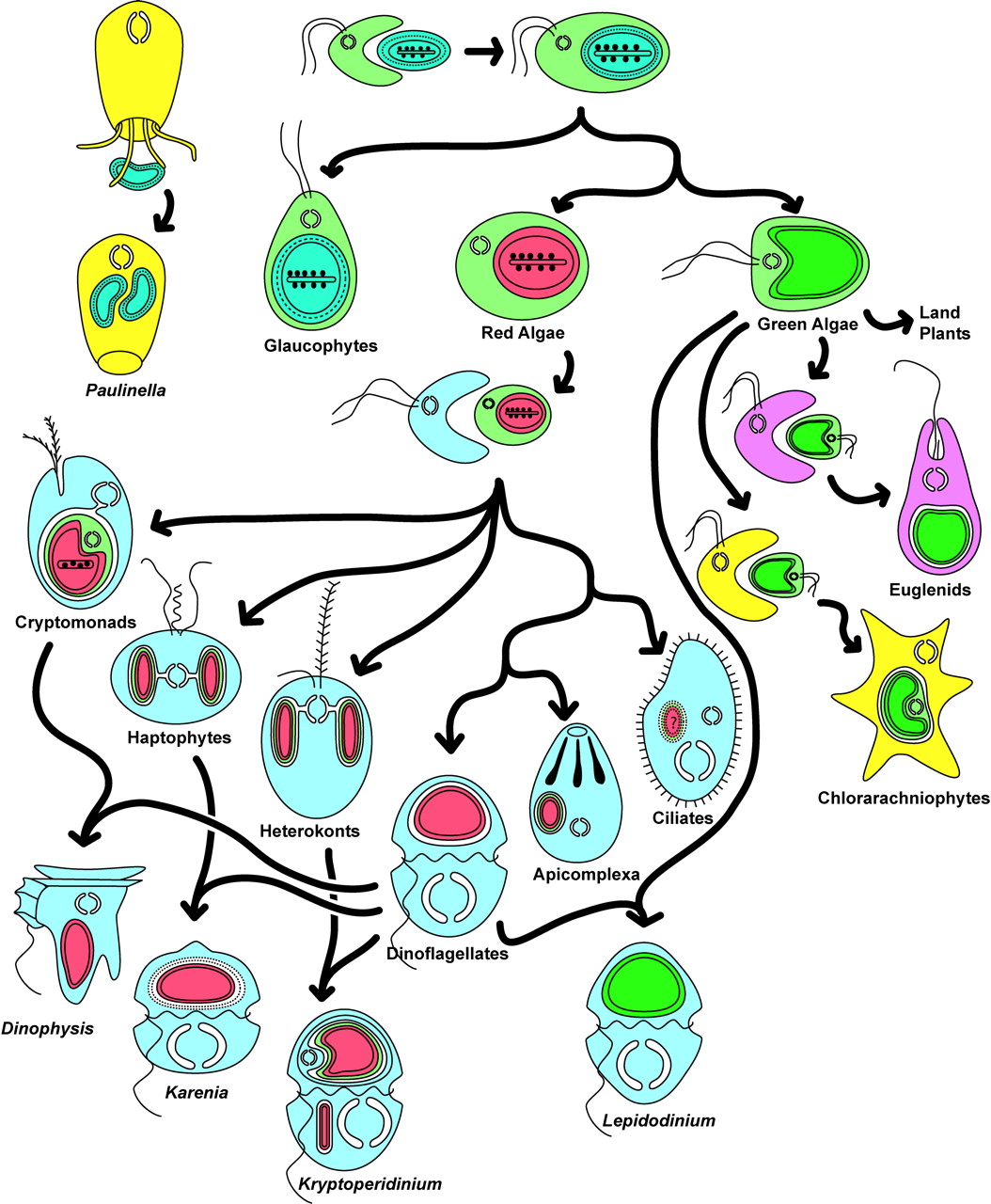
Far from being confined to the distant past, these processes continue to be observed. Certain single-celled algae, including cryptophytesSingle-celled organisms, most of which are photosynthetic. Their chloroplasts are enclosed by four membranes, indicating the integration of a red alga through secondary endosymbiosis. The presence of a nucleomorph in many species makes them an important model for understanding the evolution of complex photosynthetic organelles. Cryptophytes are found in many habitats, particularly aquatic ones (oceanic environments, freshwater, and interstitial waters in terrestrial wetlands). Some species have become intestinal parasites of metazoans. A few are endosymbionts of dinoflagellates. and stramenopiles (heterokontsA major lineage of eukaryotes comprising both photosynthetic organisms (diatoms, brown algae, chrysophytes, etc.) and non-photosynthetic organisms (oomycetes). The photosynthetic species acquired their chloroplasts through secondary endosymbiosis with a red alga, illustrating the importance of endosymbiosis in the evolution of eukaryotes.), whose four-membraned plastids originated from a secondary endosymbiosis, can themselves occur as endosymbionts within dinoflagellates that have lost their own plastids. These associations illustrate a succession of endosymbiotic events throughout evolution and demonstrate that the integration of new cellular partners remains an ongoing evolutionary process.
Advances in phylogenomics, microbiology, cell biology and microscopy continue to refine our understanding of the origin of eukaryotes. Whilst the general principle of an endosymbiotic origin for the eukaryotic cell is now widely accepted, several questions remain unanswered, notably the origin of the nucleus, the chronology of events leading to LECA, and the precise mechanisms of α-proteobacterial internalisation. Every new Asgard archaeon discovered, every genome sequenced and every successful culture adds a new piece to this evolutionary jigsaw.
Long regarded as a biological curiosity, endosymbiosis is now recognised as one of the main drivers of evolutionary innovation. By permanently uniting previously independent organisms, it has enabled the emergence of more complex cells, contributed to the evolution of multicellularity and paved the way for the diversification of plants, fungi and animals. The history of the eukaryotic cell thus illustrates one of the greatest transitions in evolution: a transition in which stable cooperation between organisms became a major source of biological novelty.
Références & notes
Image de couverture : [Source Photo © Jacques Joyard]
[1] Figure 2 is based on data from the following references: Lang T. et al. (2000) Autophagy and the CVT pathway both depend on AUT9. J Bacteriol 182, 2125–2133. doi: ttps://doi.org/10.1128/jb.182.8.2125-2133.2000; Spang, A., Saw, J. H., Jørgensen, S. L., Zaremba-Niedzwiedzka, K., et al. (2015). Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature, 521(7551), 173–179. https://doi.org/10.1038/nature14447; Zaremba-Niedzwiedzka, K., Caceres, E. F., Saw, J. H., Bäckström, D., Juzokaite, L., Vancaester, E., … Ettema, T. J. G. (2017). Asgard archaea shed light on the origin of eukaryotic cellular complexity. Nature, 541(7637), 353–358. https://doi.org/10.1038/nature21031; Imachi, H., Nobu, M. K., Nakahara, N., Morono, Y., Ogawara, M., Takaki, Y., … Takai, K. (2020). Isolation of an archaeon at the prokaryote–eukaryote interface. Nature, 577(7791), 519–525. https://doi.org/10.1038/s41586-019-1916-6
[2] Selosse, M.A. (2012). The rise and fall of the endosymbiotic theory. La Recherche 468: 92–94.
[3] Zaremba-Niedzwiedzka, K. et al. (2017). Asgard archaea shed light on the origin of eukaryotic cellular complexity. Nature, 541(7637), 353–358. https://doi.org/10.1038/nature21031
[4] Tobiasson, V., Luo, J., Wolf, Y.I. et al. (2026). Dominant contribution of Asgard archaea to eukaryogenesis. Nature 650, 141–149.
[5] Zhang, J., Feng, X., Li, M. et al. (2025). Deep origin of eukaryotes outside Heimdallarchaeia within Asgardarchaeota. Nature 642, 990–998 https://doi.org/10.1038/s41586-025-08955-7
[6] Richards T.A., Eme L., Archibald J.M., Leonard G., Coelho S.M., de Mendoza A. et al. (2024) Reconstructing the last common ancestor of all eukaryotes. PLoS Biology 22(11): e3002917. https://doi.org/10.1371/journal.pbio.3002917
[7] Schleper, C. & Rodrigues-Oliveira, T. (2026) Asgard archaea: have we found our microbial ancestors? EMBO J 45, 1836–1851.
[8] Martin W.F., Garg S. & Zimorski V. (2015) Endosymbiotic theories for the origin of eukaryotes. Phil. Trans. R. Soc. B 370, 20140330. doi: 10.1098/rstb.2014.0330.
[9] Martin W. & Müller M. 1998 The hydrogen hypothesis for the first eukaryote. Nature 392, 37–41.
[10] López-García P & Moreira D. (2019) Eukaryogenesis, a syntrophy affair. Nat Microbiol. 4(7):1068–1070. doi: 10.1038/s41564-019-0495-5.
[11] Imachi, H., Nobu, M.K., Nakahara, N. et al. (2020) Isolation of an archaeon at the prokaryote–eukaryote interface. Nature 577, 519–525. https://doi.org/10.1038/s41586-019-1916-6
[12] Selosse M.A., Albert B. & Godelle B. (2001) Small is successful: selection for reducing organelle genome size favours gene transfer to the nucleus. Trends Ecol Evol 16, 135–141.
[13] Tobiasson, V., Luo, J., Wolf, Y.I. et al. (2026) Dominant contribution of Asgard archaea to eukaryogenesis. Nature 650, 141–149. https://doi.org/10.1038/s41586-025-09960-6.
[14] Archibald JM & Keeling PJ (2002) Recycled plastids: a ‘green movement’ in eukaryotic evolution. Trends in Genetics 18, 577–584.
[15] Keeling PJ (2004) Diversity and evolutionary history of plastids and their hosts. Am. J. Bot. 91, 1481–1493.
[16] Douce R., Block M.A., Dorne A.J., Joyard J. (1984) The plastid envelope membranes: their structure, composition, and role in chloroplast biogenesis. Subcell. Biochem. 10, 1–84, Springer US (Ed.)
[17] Cruz S, Cartaxana P (2022) Kleptoplasty: Getting away with stolen chloroplasts. PLoS Biol 20(11): e3001857. https://doi.org/10.1371/journal.pbio.3001857
[18] Selosse MA & Loiseaux-de Goër S (1997) The Saga of Endosymbiosis, La Recherche 296, 36
[19] Stairs C.W., Leger M.M., Roger A.J. (2015) Diversity and origins of anaerobic metabolism in mitochondria and related organelles. Philos. Trans. R. Soc. B 370, 20140326; see also Stechmann A. et al. (2008) ‘Organelles in Blastocystis that blur the distinction between mitochondria and hydrogenosomes’. Curr. Biol. 18, 580–585.
[20] Lefèvre T., Renaud F., Selosse M.-A., Thomas F. (2010). Evolution of interactions between species, in F. Thomas, T. Lefèvre & M. Raymond (eds.), Evolutionary Biology, pp. 530–613. De Boeck, Paris.
[21] Keeling PJ (2010) The endosymbiotic origin, diversification and fate of plastids. Phil. Trans. R. Soc. B 365, 729–748
[22] Marin B., Nowack E.C.M., Melkonian M. (2005) A plastid in the making: evidence for a second primary endosymbiosis. Protist 156, 425–432; see also Delaye L., Valadez-Cano C., Pérez-Zamorano B. (2016) How really ancient is Paulinella chromatophora? PLoS Curr. Tree of Life 8.
[23] Molina J., Hazzouri K.M., Nickrent D. et al. (2014), Gisela P. Concepcion, Michael D. Purugganan, ‘Possible Loss of the Chloroplast Genome in the Parasitic Flowering Plant Rafflesia lagascae (Rafflesiaceae)’, Molecular Biology and Evolution, 31, 4, pp. 793–803, https://doi.org/10.1093/molbev/msu051
[24] Glaucophytes: Single-celled organisms with flagella (two of unequal length) living in freshwater pools in temperate regions.
[25] Selosse MA, Albert B and Godelle B (2001) Small is successful: selection for reducing organelle genome size favours gene transfer to the nucleus. Trends in Ecology and Evolution 16, 135–141.
[26] Jarvis P (2004) Organellar proteomics: Chloroplasts in the spotlight. Current Biology 14, R317–9. http://www.cell.com/current-biology/references/S0960-9822%2804%2900231-3
The Encyclopedia of the Environment by the Association des Encyclopédies de l'Environnement et de l'Énergie (www.a3e.fr), contractually linked to the University of Grenoble Alpes and Grenoble INP, and sponsored by the French Academy of Sciences.
To cite this article: SELOSSE Marc-André, JOYARD Jacques (July 20, 2026), Symbiosis and evolution: at the origin of the eukaryotic cell, Encyclopedia of the Environment, Accessed July 29, 2026 [online ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/en/life/symbiosis-and-evolution-origin-eukaryotic-cell-2/.
The articles in the Encyclopedia of the Environment are made available under the terms of the Creative Commons BY-NC-SA license, which authorizes reproduction subject to: citing the source, not making commercial use of them, sharing identical initial conditions, reproducing at each reuse or distribution the mention of this Creative Commons BY-NC-SA license.


