




气候变化和全球化,昆虫入侵的驱动力

全球变暖和贸易全球化对昆虫的地理分布具有决定性的影响。随着商业网络的发展,世界各地非本土昆虫种群建立的情况呈指数增长。同时,全球变暖正促使许多昆虫向北扩散到更高纬度的地区。这两个因素共同解释了近年出现的多次昆虫入侵现象。
1. 欧洲非本地昆虫种群建立的爆炸式增长
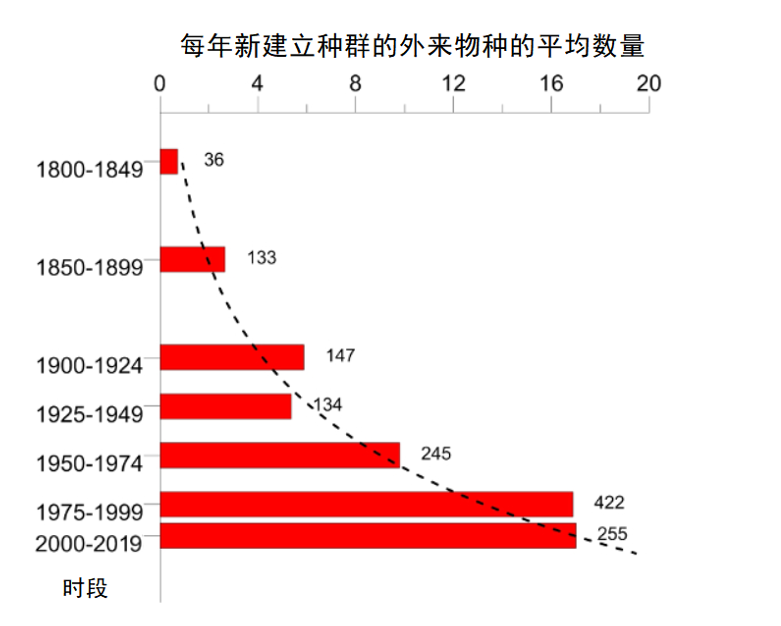
当满足以下条件时,可认定一个物种构成入侵:(a)从外界(其发源地大陆以外)引进,(b)在本地建立种群(成功繁殖),然后(c)扩散至登陆地之外,并造成(d)一系列负面影响(经济、健康、社会),破坏当地生态系统的稳定。
尽管自然栖息地的破坏是目前生物多样性下降的主要原因,但非本地物种入侵已经成为第二大威胁。自20世纪初以来,非本地昆虫(即被引进至其发源地以外的大陆的昆虫)建立种群的速度呈指数增长。在欧洲新建立种群的物种数量从1950-1974年期间的每年9.7个,增至1990-2016年期间的17.0个[1](图1)。到2016年底,确定引进并于欧洲建立种群的非本地昆虫物种达1418种[2],使其成为仅次于植物的第二大类非本地物种[3](参阅“当入侵植物也在田间蔓延”)。在其他大陆上也出现了类似的情况[4]。

这种传播速度的加快在很大程度上(> 90%)是由与人类活动有关的意外介入引起,并且主要与全球观赏植物贸易的快速增长有关,因为这让植物相关物种得以在全球运输[5]。然而,也有物种的传播与特定的生物学原因无关,例如通过容器或货物被引入,例如亚洲虎蚊(白纹伊蚊)或亚洲墨胸胡蜂(图2)。
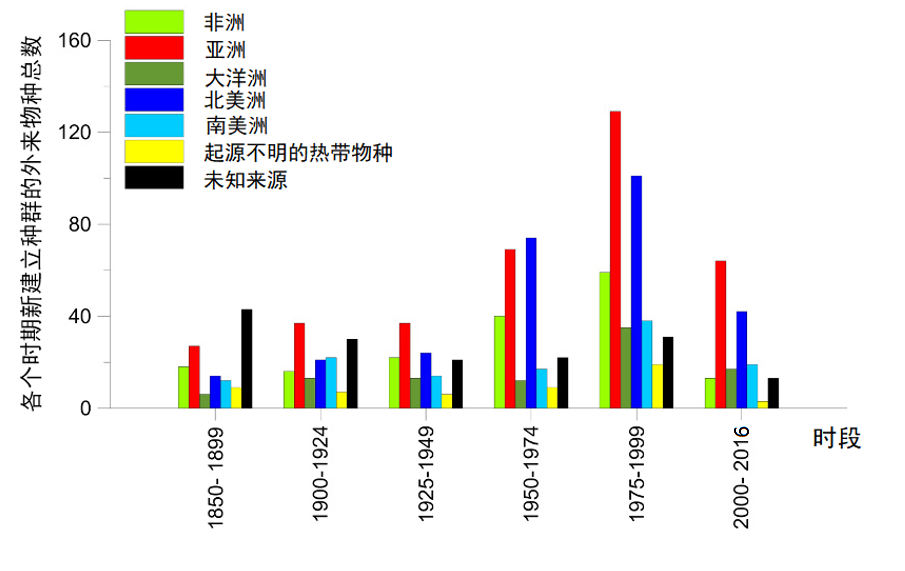
当前,非本地昆虫物种的数量一直保持持续稳定的增加[4]。而且由于新市场、新销路的拓展,以及运输货物本身的特性,昆虫的输入来源区域显著扩大。因此,亚洲成为欧洲入侵昆虫物种的新的主要来源地(>30%),取代了此前的主要来源地北美(图3)。
如今,(某些)定性为“新兴”的物种(称为“新兴”的原因是它们此前从未被引入至起源地以外的大陆[6])已成为入侵地的优势物种。例如2004年在法国发现的亚洲大黄蜂就是这种情况。
2. 非本地物种在欧洲的传播速度越来越快
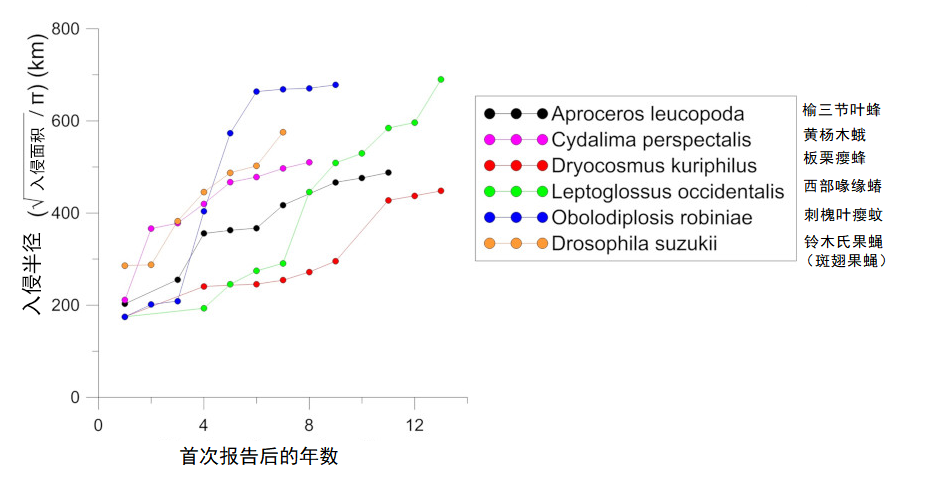
非本地昆虫扩张速度正在加快。过去二十年,在欧洲建立种群的非本地物种的平均扩张速度比起二战后刚进入时明显加快[1]。在不到十五年的时间里,许多入侵物种已几乎遍布整个欧洲大陆,然而,这在20世纪上半叶要花费数十年才能完成。部分案例表明,这种扩张速度与所入侵昆虫的来源地并不相关(图4和5)。

来自北美原生地的西部喙缘蝽(Leptoglossus occidentalis)(图4),于1999年在意大利东北部首次发现[7]。在2018年,它便扩散到葡萄牙、俄罗斯、从北非到斯堪的纳维亚半岛,以及中东、亚洲和南美洲。
亚洲起源的昆虫传播速度甚至更快,以不到10年的时间席卷了欧洲大部分地区,这些物种有:
- 黄杨木蛾(Cydalima perspectalis)(图4)最早于2006年在德国发现[8]。
- 斑翅果蝇(Drosophila suzukii)(图4)于2008年在意大利发现[9]。对于起源于南美的番茄潜叶蛾(Tuta absoluta)(图4)也是如此,最早于2006年在西班牙发现[10]。
对过去250年中来到欧洲的1171种非本地昆虫进行分析表明,1990年后引入物种的初始扩张速率:
- 比第二次世界大战(1950-1969)后建立种群的物种高4倍,
- 比1970-1989期间发现的物种高3倍[1]。
近期非本地昆虫扩张速度的加快(图5)可能与欧洲的政治和经济变化(柏林墙倒塌和欧盟的扩大)有关,尤其是欧盟内部的海关管制的取消[1]。
许多昆虫可自然传播,有时传播的距离很远,但由于入侵区域是逐年扩展的,对大多数被研究的物种而言,将自然传播作为合理解释仍比较难令人信服。例如:
- 黄杨木蛾(Cydalima perspectalis)和西部喙缘蝽(Leptoglossus occidentalis)在一年内扩散半径超过150 km(图4和5),而其成虫飞行所能到达的最远距离估计只有几十公里[8]。
- 一种起源于北美的小型蚊虫刺槐叶瘿蚊(Obolodiplosis robiniae)(成虫尺寸:5毫米),两年内扩张范围超过450公里(图5和6)[1]。

对于部分物种而言,入侵地区和原生地区之间的种群遗传多样性比较表明:存在同一种昆虫不同原生地区的多源引入的情况。也可能涉及“桥头堡(Bridgehead)”现象,即引入的昆虫来自于已入侵的地区[7]。这种多源引入可解释某些物种的快速传播,例如黄杨木蛾[8]、西部喙缘蝽[7]和亚洲长角甲虫(光肩星天牛,Anoplophora glabripennis)[11](图7)。然而,遗传分析也表明:人类活动,特别是植物贸易,对这些非本地昆虫在欧洲的快速传播起着关键的作用。黄杨木蛾便是一个典型案例:其卵和毛虫通过苗圃和花卉中心的观赏黄杨木贸易传播至整个欧洲[8](图4)。
3. 全球变暖正在改变昆虫的分布范围
根据政府间气候变化专门委员会(IPCC [12])的模型,自20世纪初以来全球平均温度升高了0.61°C,2100年时变暖导致的升温将达到2.6至4.8°C之间,由此可以预见气候变化将对大多数昆虫物种产生显著影响。这种可预见的影响与昆虫的生物学特性直接相关。
昆虫是不产生热量的“冷血”动物。与“内温型(endothermic)”哺乳动物通过新陈代谢产生热量并保持体温恒定不同,昆虫被称为“外温动物”,其体温和有关生理过程直接取决于环境温度的变化。这些变化与其他因素一起,共同决定它们的活动周期和节奏、幼虫发育的不同阶段所持续的时间和年世代数,以及它们的飞行可能性等。对于许多物种,特定的温度阈值会解除其地理分布的限制:
- 由此,-16°C等温线限制了北美南部松小蠹(一类针叶树种的主要害虫,学名Dendroctonus frontalis [13])的冬季存活可能性(图8),例如松异舟蛾的幼虫(参阅“松异舟蛾:全球变暖影响的典型”)
- 反之,+40°等温线也限制了这种飞蛾的卵在夏季的存活[14]。

- 过去一个世纪,欧洲变暖约0.8°C,平均等温线向极点移动了120 km[15],与此同时,63%的非迁移性蝴蝶分布区域向北延伸了35~240 km的距离。
- 在北美西部,斑蝶(Euphydryas editha)分布区域平均向北扩张92公里,这也与同期等温线向北位移105km有关[16]。
- 在过去的30年中,已观察到甲虫、蜻蜓和蚱蜢有类似的分布区域向北和更高海拔移动的现象[17]。
但是,将物种过去和当前的分布与同时期等温线的移动进行简单比较,并不足以证明气候变化的影响。为了证明这不是简单相关性造成的,我们必须确定分布范围变化与所考虑的气候因素变化之间的因果关系。

在温带地区,低温通常是限制昆虫物种分布范围的关键因素。昆虫冬季的生存以及发育阶段的转化(例如卵、幼虫、若虫和成虫)都存在最低温度阈值。因此,冬季温度略微的升高即会使其能在原先因恶劣的气候条件无法生存的地区存活:
- 南松大小蠹(Dendroctonus frontalis-图8)冬季致死温度约为-16°C,本世纪初之前只能在美国东南部生存。该区域当时是昆虫分布的北部极限。-16°C等温线沿北纬方向的移动,使得该昆虫在美国东北部更北的区域定居[18]。
- 北欧斯堪的纳维亚北部以前因冬季温度不适合几何纹理蝴蝶(geometric butterflies)卵的存活,现在这种情况也发生了改变(图9)。
- 针对冬季变暖在松异舟蛾扩张中起到的作用,已经有实验证明两者之间存在因果关系(Cause-effect relationships )(参阅“松异舟蛾:全球变暖影响的典型”)[19]。
4. 年平均气温的升高,是与之有关的变量吗?
全球变暖最常用的评估方法是根据海平面平均气温的升高进行评估。然而,昆虫种群并不是依据平均温度,而是依据其生活环境所处天气条件下的每日变化(温度)来进行评估。仅考虑年度气温升高不够全面,不足以探明季节性变暖(造成)的不同影响。
因此,春、夏、秋、冬四季的升温在种群扩大与否上所造成的生物学影响并不相同。冬季温度的升高使某些物种能够在先前不利的地区生存,春季和/或夏季温度的升高将可能加速其幼年阶段的发展,从而使某些物种的年世代数增加,进而为其迁移创造出有利条件。
- 对于迁徙性的蝴蝶物种尤其如此,例如夜蛾Autographa gamma,温度升高帮助它们从北非的越冬点入侵至英国[20]。
- 在意大利阿尔卑斯山,2003年夏季的热浪也促使松异舟蛾扩展其分布范围。因为夜晚温度超过了雌蛾飞行所需要阈值(14°C)的天数达到了正常值的5倍,这也导致了更多的飞蛾散布到更远的地区[21]。

然而,年温度升高也可能掩盖了季节性变暖所造成的相反作用(contradictory effects),例如日本的稻绿蝽(Nezara viridula)(图10)。这种虫子能否向北扩张,取决于全球变暖正面和负面效应是否达到平衡:冬天成虫存活率提高,秋季加速幼虫发育是正向效应,但夏季对幼虫发育会产生负面影响。同样,尽管2002-2003年显著的冬季变暖使松异舟蛾幼虫能够在巴黎地区扩张和生存,但2003年夏季的热浪使这个扩张的种群消失了很大一部分[13]。因此,对于未来频率还会有所上升的极端事件(例如夏季热浪),其额外影响也应予以考虑,尽管目前我们对此知之甚少[17]。
5. 北扩南缩?
很少有数据表明随着温度的升高,昆虫的南端分布界限区域会收缩。大多数的欧洲蝴蝶中,向北移动的物种的南端分布界限似乎仍保持稳定,只有极少数,例如灰蝶(Heodes tytirus),在南部地区的数量减少[15]。基于未来可预见的升温模拟表明,对于两种蛾(舞毒蛾Lymantria dispar和模毒蛾L. monacha),由于过高温度在冬季滞育(diapause)(一种因应不利的气候条件而暂停发育的现象)过程中对卵孵化为幼虫产生负面影响[22],使其南端分布界限缩回100到900 km的距离。
一般而言,对于在春季和夏季发育的昆虫,其分布范围的变化大致由发育速度(快或慢)和冬季滞育状态共同决定[23]:
- 对于没有滞育过程的快速发育的物种,其世代数会增加,并且分布范围扩大(例如蚜虫物种);
- 对于快速成长,但存在受温度影响的滞育过程的物种,其分布面积会缩小(例如欧洲孔雀蛱蝶,Inachis io或小帝王蛾,Saturnia pavonia);
- 对于受温度影响且发育缓慢的滞育物种,其数量将趋于减少且分布面积也将减小(例如栎枯叶蛾,Lasiocampa quercus)。
6. 全球化与全球变暖协同作用
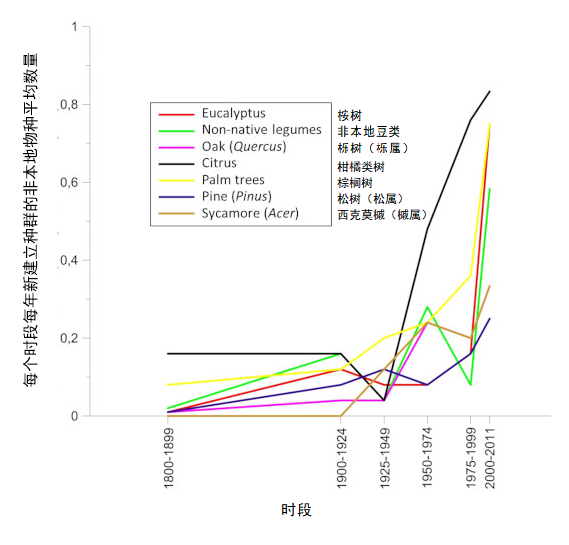
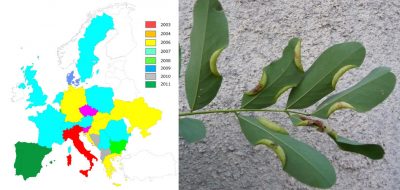
贸易的全球化促使亚热带或热带地区的昆虫物种意外来到欧洲大陆。然而,直到最近至今它们的种群建立仍受到以下限制:冬季天气不利于其在大陆大部分地区生存,利于其发育的温度条件持续时间有限。但是,自20世纪90年代以来,在欧洲种植的热带或亚热带起源的植物(如桉树,棕榈树和其他合欢树(acacias))上,非本地昆虫的种群建立相较过去有明显增长[24](图11)。到本世纪00年代末,来自亚热带或热带气候区在欧洲建立种群的昆虫已有400多种[25]。这些物种分布通常仅限在引进地,常在地中海地区,但是近年来,与棕榈树有关的昆虫入侵现象时有发生(图12),也进一步说明这些物种的分布情况也有所改变。

虽然直到21世纪中期,红棕象甲(Rhynchophorus ferrugineus)和棕榈蝶蛾(Paysandisia archon butterfly)一直局限分布于在西班牙南部和加那利群岛的传入地带。但在2004年至2007年期间(图12),它们就占领了地中海大部分地区,棕榈蝶蛾的分布区域甚至还远至英格兰南部。之所以会出现这种迅速的蔓延,是因为全球变暖产生了综合影响,让这些热带物种得以更好地发育,而观赏性棕榈的贸易发展也是一大原因。随着用于装饰雅典奥运会(2004年)的西班牙棕榈树的进口,红棕象甲也随之到来[26]。类似情况还有本地昆虫由人类意外地运输至超出其自然分布范围以外的地区。这些引入地区新的气候条件使它们得以在本地扎根,特别是那些被向北运输的地中海昆虫,例如欧洲螳螂(薄翅螳螂,Mantis religiosa)[27]或松异舟蛾(Thaumetopoea pityocampa,参阅“松异舟蛾:全球变暖影响的典型”)。
7. 只有全球变暖造成了昆虫扩散吗?
迄今为止,大多数研究都集中在升温的影响上。但是,气候变化还包含许多其他因素,例如湿度、降雨强度和频率、辐射或温室气体水平,这些因素可以直接发挥作用,也可以与升温产生协同作用。例如,在我们(法国)地区,亚洲虎蚊(白纹伊蚊,Aedes albopictus)的种群建立受到温度的影响,但同样还受到昼夜长短(光周期)、湿度和降雨的限制[28]。但是其他一些在实验室条件下进行的研究,像昆虫对二氧化碳等各种变量波动的可能响应等,通常会得出相互矛盾的结果[29]。因此,开展大规模野外实验显得尤为必要。
关于气候变化和昆虫扩散之间关系的研究,通常集中在对经济或健康有重大影响的昆虫的具体响应上,很少考虑与其相关的其他生物。然而,气候变化会同时影响整个生态系统,包括宿主(植物)、竞争性昆虫和天敌。对于相同程度的升温,(生态系统的)不同组分的响应可能会有所不同,并且相互的作用可能会在一个方向或另一个方向上发生显著变化。例如研究表明:某些捕食者 (predators)的捕食效力(effectiveness)可能会增强,例如瓢虫在温暖的环境中可能会增加(效力),蚜虫(猎物)的扩张可能会减慢[30]。另一方面,松异舟蛾比其自然天敌向北扩张得更快[31]。未来,开展同时考虑生态系统所有组分的整合研究非常必要。
8. 结论
- 世界贸易,尤其是观赏植物贸易的加强和多样化,导致了欧洲非本地昆虫数量的增加。
- 这些昆虫建立种群后,在欧洲的传播速度往往比在上世纪快。
- 气候变暖,特别是冬季温度上升,消除了限制许多本地或非本地物种活动范围的气候障碍,使这些物种扩展到以前不利于其在冬季生存的地区。
- 变暖促进了起源于亚热带和热带的非本地物种的种群建立和传播。
参考资料及说明
封面图片:2015-2016年冬季在意大利皮埃蒙特,阿尔卑斯山乌尔克斯(Italian Alps. Oulx, Piedmont)松异舟蛾沿纵向扩张的大规模攻击。 [来源:Photo Roques © INRA]
[1] Roques A., Auger-Rozenberg M.-A., Blackburn T.M., Garnas J.R., Pyšek P. et al. (2016) Temporal and interspecific variation in rates of spread for insect species invading Europe during the last 200 years. Biological Invasions 18, 907-920.
[2] Source: DAISIE European project database (“Delivering Alien Invasive Species Inventories in Europe”; http://www.europe-aliens.org) recently updated in the EASIN database (“European Alien Species Information Network“; http://easin.jrc.ec.europa),
[3] DAISIE (2009) Handbook of alien species in Europe. Berlin: Springer Science + Business Media B.V.
[4] Seebens H., Blackburn T.M., Dyer E.E., Genovesi P., Hulme P.E. et al. (2017) No saturation in the accumulation of alien species worldwide. Nature Communications. DOI: 10.1038/ncomms14435.
[5] Roques A., Rasplus J.Y., Rabistch W., Lopez-Vaamonde C., Kenis M et al. (2010). Alien Terrestrial Arthropods of Europe. BioRisk 4 (1 and 2), 1-1024.
[6] Seebens A., Blackburn T.M., Dyer E.E., Genovesi P., Hulme P.E. et al. (2018). The global rise in emerging alien species results from increased accessibility of new source pools. Proceedings of the National Academy of Sciences of the USA. doi/10.1073/pnas.1719429115.
[7] Lesieur V., Lombaert E., Guillemaud T., Courtial B., Strong W. et al. (2018) The rapid spread of Leptoglossus occidentalis in Europe: a bridgehead Invasion. Journal of Pest Science. https://doi.org/10.1007/s10340-018-0993-x
[8] Bras A. Invasion fulgurante de la pyrale du buis en Europe: Caractérisation de la diversité génétique de Cydalima perspectalis (Walker, 1859) et approche phylogéographique https://belinra.inra.fr/doc_num.php?explnum_id=1021 (in french)
[9] Cini A., Ioriatti C., Anfora G. (2012). A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bulletin of Insectology 65(1) 149-160.
[10] Desneux N., Luna M.G., Guillemaud T. and Urbaneja A. (2011) The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond the new threat to tomato world production. Journal of Pest Science 84, 403 – 408.
[11] Javal M., Roques A., Haran J., Herard F., Keena M., Roux G. (2017) Complex invasion history of the Asian long-horned beetle: fifteen years after first detection in Europe. Journal of Pest Science DOI 10.1007/s10340-017-0917-1.
[12] http://ar5-syr.ipcc.ch/
[13] Ungererer M.J., Ayres M.P., Lombardero M.J. (1999). Climate and the northern distribution limits of Dendroctonus frontalis Zimmermann (Coleoptera: Scolytidae). Journal of Biogeography 26, 1133-45.
[14] Faucet C., Rousselet J., Pineau P.; Miard F., Roques A. (2013). Are heat waves susceptible to mitigate the expansion of a species progressing with global warming? Ecology and Evolution 3(9), 2947-57.
[15] Parmesan C, Ryrholm N, Stefanescu C et al (1999). Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 399, 579-83.
[16] Parmesan C (1996). Climate change and species’ range. Nature 382, 765-766.
[17] Hickling R., Roy D.B., Hill J.K., Fox R., Thomas C.D. (2006). The distributions of a wide range of taxonomic groups are expanding polewards. Global Change Biology 12, 450- 455.
[18] Trân J.K., Ylioja T., Billings R.F., Régnière J., Ayres M.P. (2007). Impact of minimum winter temperatures on the population dynamics of Dendroctonus frontalis. Ecological Applications 17, 882-899.
[19] Jepsen J.U., Hagen S.B., Ims R.A., Yoccoz N.G. (2008). Climate change and outbreaks of the geometrids Operophtera brumata and Epirrita autumnata in subarctic birch forest: evidence of a recent outbreak range expansion. Journal of Animal Ecology 77, 257-264.
[20] Chapman J.W., Reynolds D.R., Smith A.D., Riley J.R., Pedgley D.E., Woiwod I.P. (2008). Wind selection and drift compensation optimize migratory pathways in a high-flying moth. Current Biology 18, 514-518.
[21] Battisti A. Stastny M., Buffo E., Larsson S. (2006). A rapid altitudinal range expansion in the pine processionary moth produced by the 2003 climatic anomaly. Global Change Biology 12, 66- 671.
[22] Vanhanen H., Veteli T.O., Pailvinen S., Kellomaki S., Niemala P. (2007). Climate change and range shifts in two insect defoliators: Gypsy moth and nun moth – A model study. Silva Fennica 41, 621-38 ;
[23] Bale J.S., Masters G.J., Hodkinson I.D. et al. (2002). Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Global Change Biology 8, 1-16.
[24] Eschen R., Roques A., Santini, A. (2015). Taxonomic dissimilarity in patterns of interception and establishment of alien arthropods, nematodes and pathogens affecting woody plants in Europe. Diversity and Distributions 21 (1), 36-45.
[25] Walther G R., Roques A., Hulme P.E., Sykes M.T., Pyšek P., et al. (2009). Alien species in a warmer world: risks and opportunities. Trends in Ecology and Evolution 24, 686-692.
[26] Roques A. (2010). Alien forest insects in a warmer world and a globalized economy: Impacts of changes in trade, tourism and climate on forest biosecurity. New Zealand Journal of Forestry 40, 77-94.
[27] Ott J., ed (2008). Monitoring climate change with dragonflies. Series Faunistica 81. Pensoft, Sofia. Cannon R.J.C. (1998). The implications of predicted climate change in the UK, with emphasis on non-indigenous species. Global Change Biology 4, 785- 796.
[28] Eritja R., Escosa R., Lucientes J. et al. (2005). Worldwide invasion of vector mosquitoes: Present European distribution and challenges for Spain. Biological Invasions 7, 87-97.
[29] Newman J.A. (2005). Climate change and the fate of cereal aphids in Southern Britain. Global Change Biology 11, 940-944.
[30] R.J.C. Cannon (1998). The implications of predicted climate change in the UK, with emphasis on non-indigenous species. Global Change Biology 4, 785-796.
[31] Auger-Rozenberg M.A., Barbaro L., Battisti A., Blache S.,. Charbonnier Y., et al. (2015). Ecological responses of parasitoids, predators and associated insect communities to the climate-driven expansion of pine processionary moth. pp. 311- 358 In Roques A. (Ed.) Processionary Moths and Climate Change: An Update. Springer/ Quae.
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: ROQUES Alain, AUGER-ROZENBERG Marie-Anne (2024年12月21日), 气候变化和全球化,昆虫入侵的驱动力, 环境百科全书,咨询于 2026年7月4日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/climate-change-globalization-drivers-of-insect-invasions-2/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。
