


Symbiose et évolution : à l’origine de la cellule eucaryote
PDF
Il y a plus de deux milliards d’années, la vie était exclusivement microbienne et occupait les océans, les sédiments marins et les environnements hydrothermaux. Dans ces milieux se sont nouées des associations étroites entre micro-organismes, dont certaines ont évolué vers de véritables endosymbioses, c’est-à-dire l’intégration durable d’un organisme à l’intérieur d’un autre. Parmi ces événements, l’un a profondément transformé l’histoire du vivant : l’association entre une archée apparentée aux actuelles archées Asgard et une α-protéobactérie, à l’origine de la mitochondrie et de la lignée eucaryote. De cette alliance sont issus les animaux, les plantes, les champignons et une immense diversité d’eucaryotes unicellulaires. La cellule eucaryote se distingue par son organisation complexe, avec un noyau renfermant l’essentiel de l’ADN, des mitochondries assurant le métabolisme énergétique et, chez les plantes et de nombreuses algues, des chloroplastes réalisant la photosynthèse. D’autres endosymbioses ont marqué l’évolution : l’acquisition d’une cyanobactérie a donné naissance aux chloroplastes, puis des endosymbioses secondaires et tertiaires ont propagé les plastes dans de nombreuses lignées. L’endosymbiose apparaît ainsi comme un moteur majeur de l’innovation évolutive et de la diversification du vivant.
1. La cellule eucaryote est une chimère
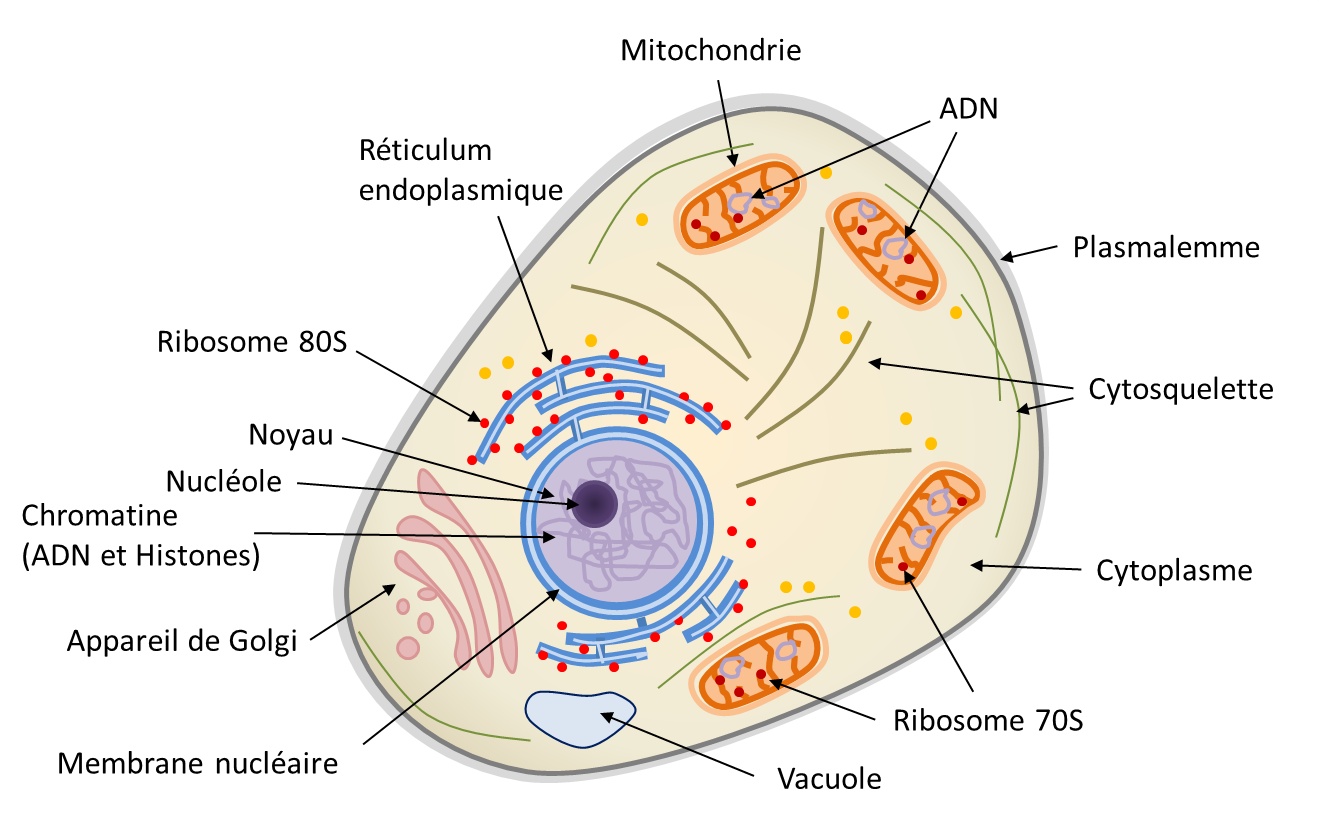
Le noyau eucaryote est délimité par une double membrane, l’enveloppe nucléaire (Figure 1). Il renferme le génome nucléaire, qui contient la très grande majorité des gènes nécessaires au fonctionnement cellulaire portés par l’ADN (acide désoxyribonucléique). Pourtant, la cellule eucaryote héberge également des génomes non nucléaires au sein de ses organites :
- le génome mitochondrial, situé dans la matrice des mitochondries (Figure 1) ;
- le génome chloroplastique, situé dans le stroma des chloroplastes (chez les plantes et les algues).
L’ADN constitutif de ces trois génomes n’est pas organisé de la même manière. Dans le noyau, le génome est réparti sur plusieurs molécules d’ADN linéaires, les chromosomes, associées à des protéines appelées histonesProtéines basiques s’associant à l’ADN pour former la structure de base de la chromatine. Les histones jouent un rôle important dans l’empaquetage et le repliement de l’ADN.. Il contient à la fois des séquences codantes (transcrites en ARN messagers puis traduites en protéines) et non codantes. La configuration tridimensionnelle du génome nucléaire est fonctionnellement cruciale : l’enroulement de l’ADN sur lui-même et autour des histonesProtéines basiques s’associant à l’ADN pour former la structure de base de la chromatine. Les histones jouent un rôle important dans l’empaquetage et le repliement de l’ADN. permet d’empaqueter une grande quantité d’information génétique dans un volume très réduit. En revanche, les génomes mitochondrial et chloroplastique sont généralement constitués de molécules d’ADN circulaires, organisées différemment de l’ADN nucléaire. Dans tous les cas, l’ADN conserve sa structure en double hélice ; seuls changent l’organisation et la forme générale de la molécule, circulaire dans les organites et généralement linéaire dans le noyau.
En effet, les cellules procaryotesMicro-organismes (en général unicellulaires) à la structure cellulaire simple, ne comportant pas de noyau, et presque jamais de compartimentation interne (la seule exception étant les thylakoïdes chez les cyanobactéries). Les procaryotes comportent deux groupes distincts, les Archées et les Bactéries. (Bactéries et ArchéesMicroorganismes unicellulaires dépourvus de noyau, présents dans presque tous les environnements, des océans et des sols aux sources hydrothermales, lacs hypersalés et autres milieux extrêmes. Les analyses phylogénétiques de Carl Woese et George E. Fox (1977) ont montré qu’elles constituent une lignée distincte des Bactéries, bien que toutes deux soient des procaryotes. Les découvertes récentes, notamment celles des archées Asgard, indiquent que les Archées sont les plus proches parentes procaryotes connues des eucaryotes et qu’elles ont joué un rôle majeur dans l’origine de la cellule eucaryote.) ne possèdent pas de noyau véritable. Leur ADN, le plus souvent circulaire, est organisé de manière similaire à celui des mitochondries et des chloroplastes, Cette organisation suggère que ces organites pourraient dériver d’anciens procaryotes. La réplication, la transcription et la traduction s’y déroulent donc directement dans le cytoplasme. Il convient toutefois de souligner que les Archées ne sont que superficiellement comparables aux Bactéries : leur métabolisme diffère profondément, et les mécanismes de réplication, transcription et traduction impliquent des protéines et des processus plus proches de ceux des eucaryotes. Enfin, les procaryotes présentent généralement une compartimentation interne limitée et moins complexe (les cyanobactéries constituant une exception notable). Surtout, leurs compartiments, lorsqu’ils existent, ne sont pas mobiles : le cytosquelette, quand il est présent, n’assure pas le déplacement organisé des composants cellulaires.
Tableau 1. Comparaison des cellules de type eucaryote et procaryote.
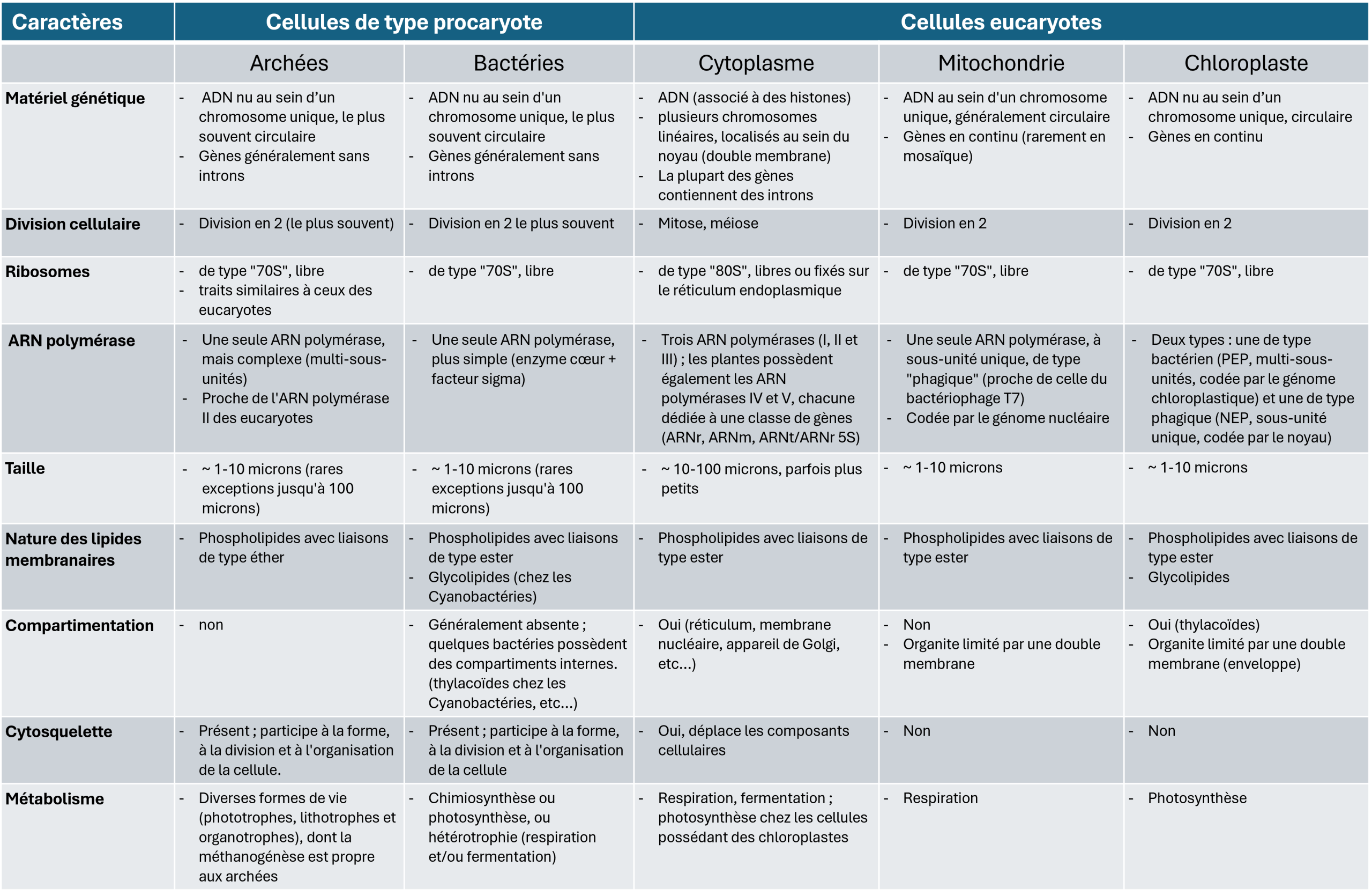
Le Tableau 1 compare les propriétés des cellules procaryotes et eucaryotes, en tenant compte des mitochondries et, le cas échéant, des chloroplastes de ces dernières. Il montre que mitochondries et chloroplastes partagent de nombreux traits avec les cellules procaryotes. Au-delà de la structure de leur ADN, les organites de la cellule eucaryote se forment à partir d’organites préexistants, ils se multiplient par fission binaire (ou scissiparité), comme les bactéries. Ils disposent aussi de la même machinerie de synthèse des protéines : des ribosomesÉnorme complexe composé d’ARN et de protéines ribosomiques, associé à une membrane (au niveau du réticulum endoplasmique granuleux) ou libre dans le cytoplasme. La fonction du ribosome est de traduire les ARN messagers (ARNm) en protéines. L’activité enzymatique du ribosome étant portée par les ARNr, le ribosome est un ribozyme. Commun à toutes les cellules (procaryotes et eucaryotes), la structure et la composition du ribosome varie selon les organismes. Chez les procaryotes et les organites cellulaires (mitochondrie, chloroplaste), le ribosome est dit 70S (S correspondant à l’unité de sédimentation dite de Svedberg) et il est constitué des sous-unités 50S et 30S. Le ribosome des eucaryotes est appelé 80S formé des deux sous-unités 60S et 40S. de type 70S, libres dans la matrice ou le stroma, alors que le cytoplasme eucaryote utilise des ribosomes de type 80S, parfois fixés sur les membranes du réticulum endoplasmiqueRéseau membranaire du cytoplasme des cellules eucaryotes, essentiel au métabolisme cellulaire (synthèses de lipides et de protéines, stockage du calcium). Associé aux ribosomes, c’est le lieu de synthèse d’une part des protéines secrétées à l’extérieur de la cellule et, d’autre part, des protéines et des lipides constituant les membranes des organites cellulaires (appareil de Golgi, lysosomes, mitochondries, noyau, ribosomes, vésicules…).. Enfin, on retrouve chez certaines bactéries le métabolisme propre aux mitochondries (la respiration) et aux chloroplastes (la photosynthèse). La cellule eucaryote se distingue en revanche par le cytosquelette, système protéique auto-organisé et mobile qui positionne et déplace les organites dans la cellule. Chez les procaryotes, un tel réseau beaucoup plus simple et beaucoup moins dynamique que celui des eucaryotes, et reste peu développé dans les mitochondries et les chloroplastes.
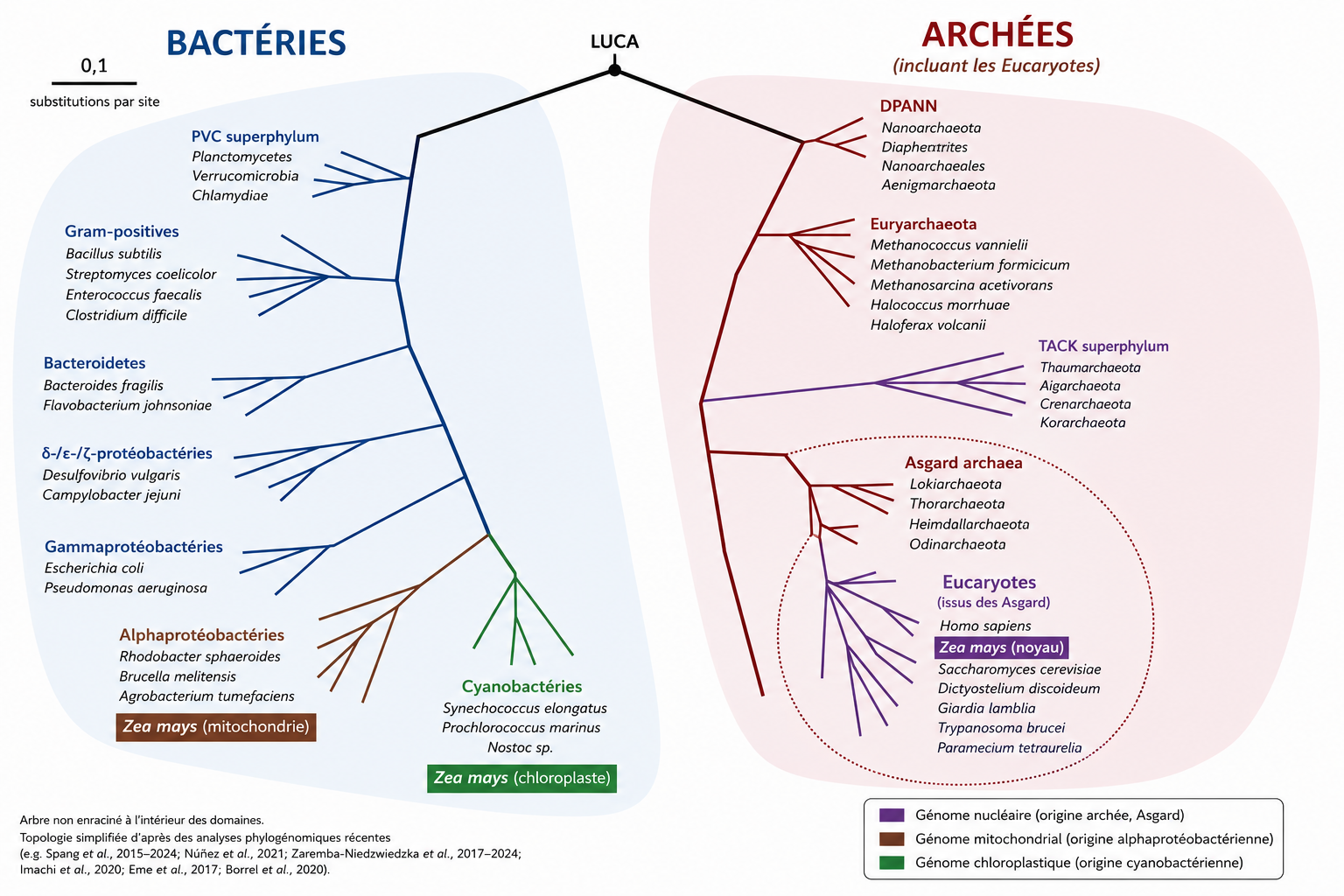
Ces propriétés montrent que la cellule eucaryote est une chimère biologique, c’est-à-dire un organisme dont le patrimoine génétique provient de plusieurs lignées évolutives distinctes : elle associe des constituants d’origine archéenne (le noyau) et des composantes d’origine bactérienne (chloroplastes, mitochondries).
Dès le tournant du 19e au 20e siècle, l’origine symbiotique des cellules eucaryotes avait déjà été pressentie mais il fallut attendre l’essor de nouvelles méthodes d’étude de la cellule — microscopie électronique, biochimie, biologie moléculaire — pour que la théorie de l’origine endosymbiotique soit remise au goût du jour, vers 1970, par la microbiologiste américaine Lynn Margulis [2] (Lire Focus Les pionniers de la théorie endosymbiotique).
Mais la découverte que les Eucaryotes émergent d’une lignée particulière d’Archées (Asgard) a révolutionné le paradigme séparant procaryotes et eucaryotes (Lire La révolution des archées Asgard).
2. Comment est apparue la cellule eucaryote ?
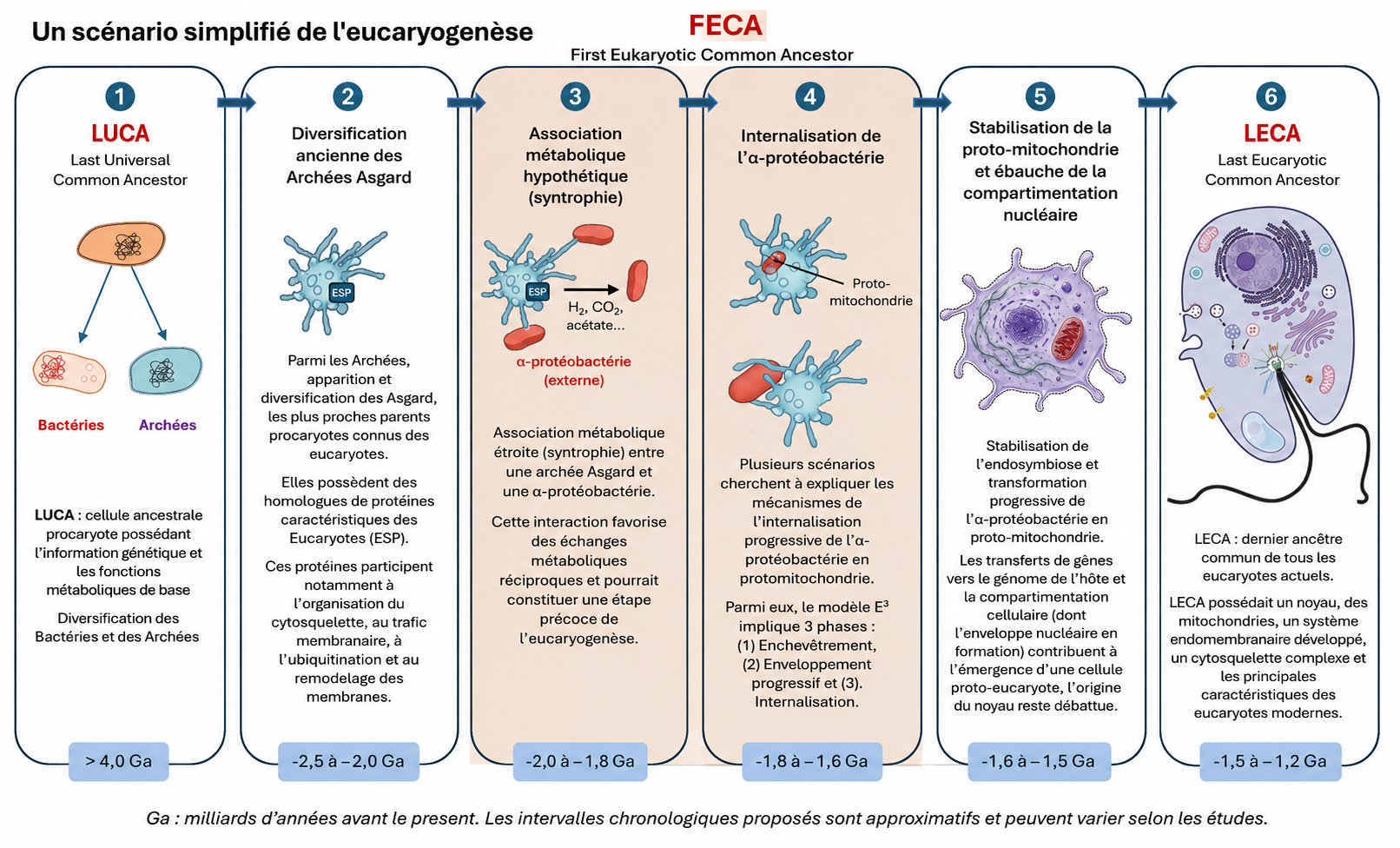
L’apparition des premières cellules eucaryotes, il y a environ 1,5 à 2 milliards d’années, représente l’une des transitions évolutives les plus importantes de l’histoire du vivant. Survenue près d’un 1,5 à 2 milliards d’années après l’émergence des premiers procaryotes (voir Focus LUCA, LECA et les ancêtres communs de l’arbre du vivant), elle a rendu possible l’émergence d’une complexité cellulaire inédite (…) et a ouvert la voie à l’évolution de la multicellularité. Les données issues de la génomique, de la phylogénomique et de la microbiologie convergent aujourd’hui vers ce scénario, qui constitue le cadre explicatif actuellement le mieux étayé pour rendre compte de l’origine des cellules eucaryotes.
2.1. La cellule hôte : une archée Asgard
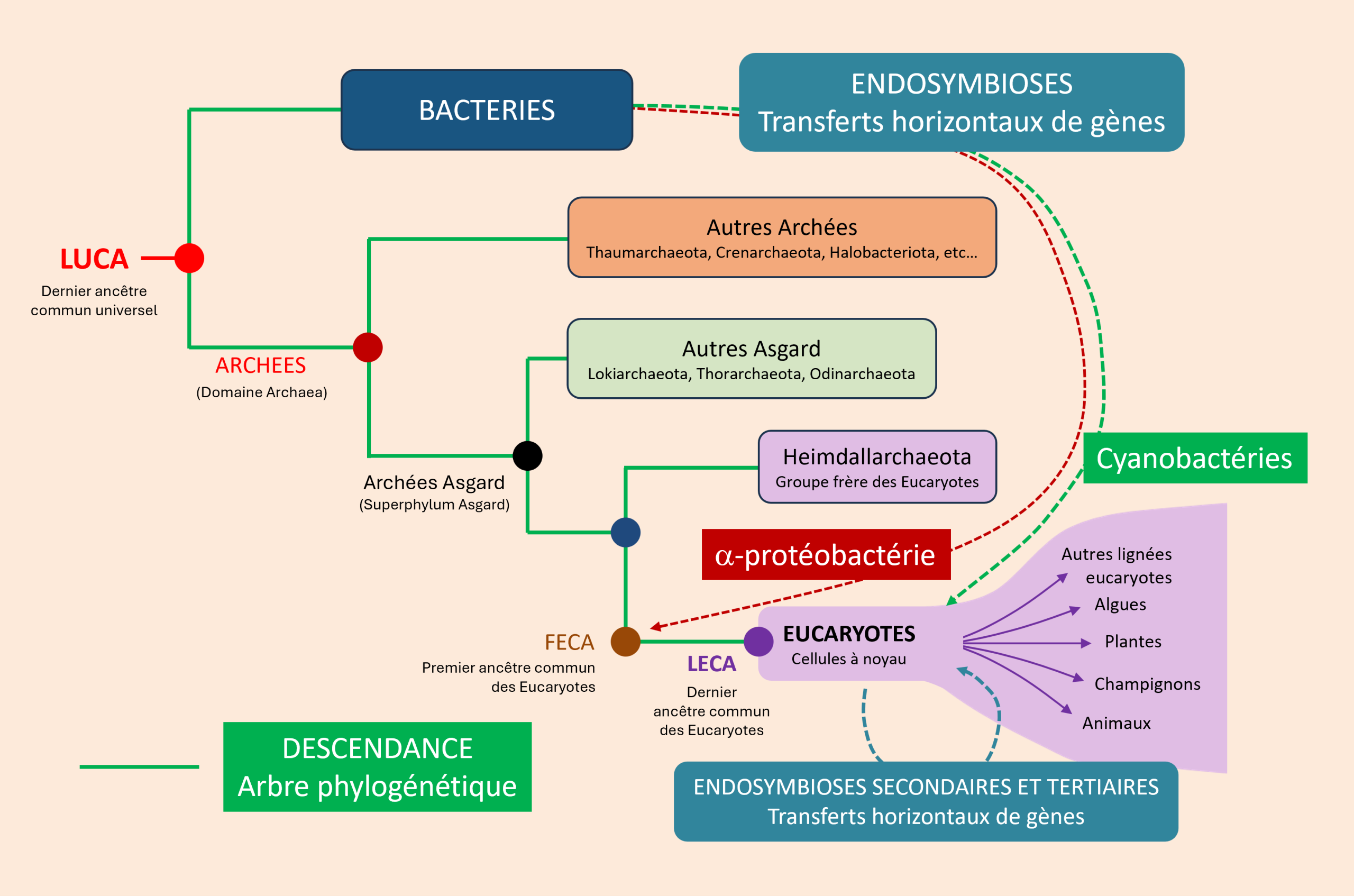
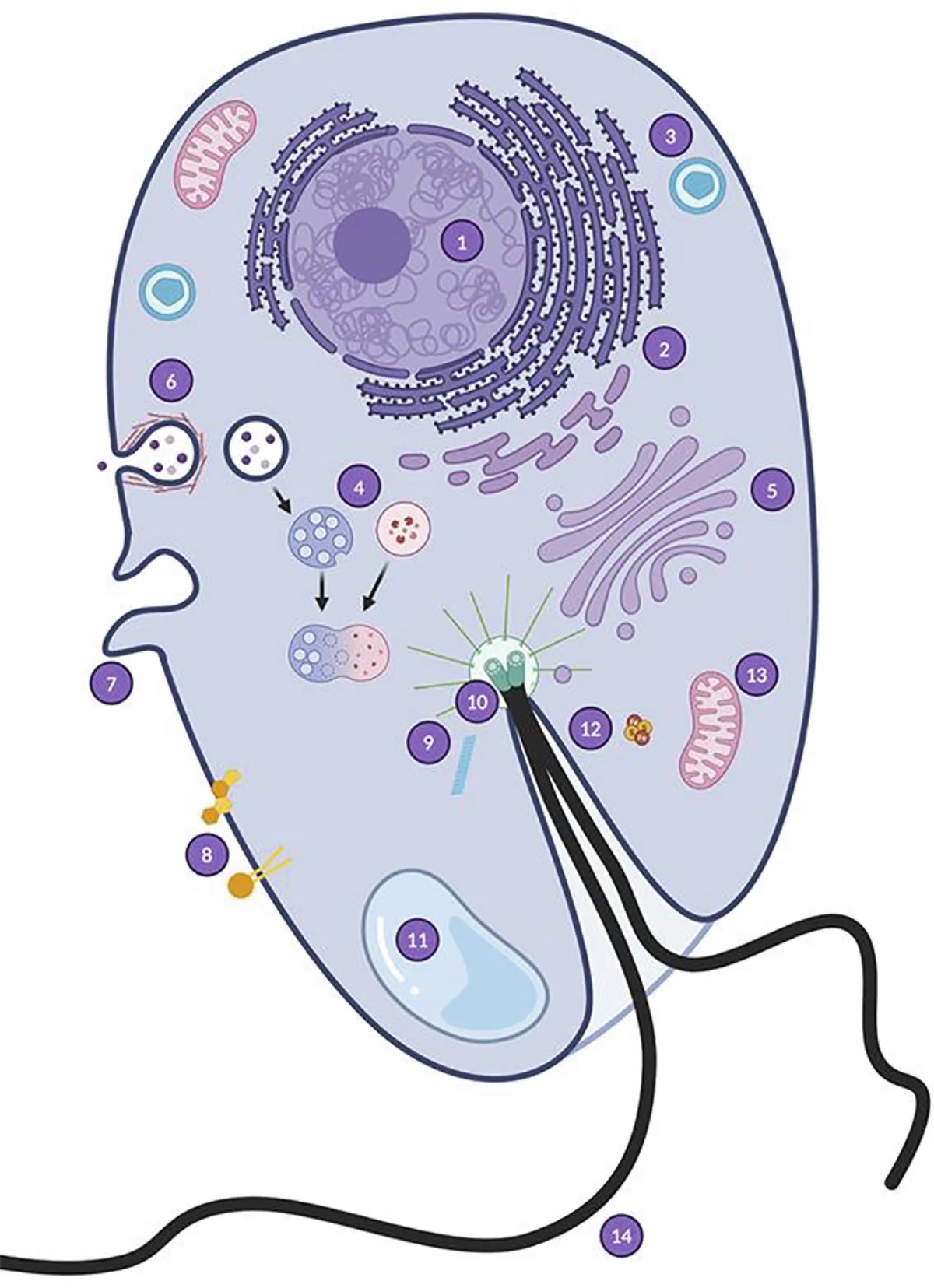
Des reconstructions récentes du dernier ancêtre commun des eucaryotes (LECA, voir Focus LUCA, LECA et les ancêtres communs de l’arbre du vivant) montrent que la grande majorité des gènes conservés d’origine non-mitochondriale proviennent d’une lignée archéenne apparentée aux Asgard actuelles, avec une contribution dominante dans la plupart des systèmes fonctionnels cellulaires (réplication, transcription, traduction, cytosquelette, endomembranes). Le LECA était déjà une cellule complexe, dotée d’un noyau, d’un cytosquelette dynamique, d’un système endomembranaire développé, de capacités de phagocytose et très probablement de sexualité (méiose) [4],[5]. La figure 4 résume schématiquement ces caractéristiques cellulaires du LECA [6].
2.2. Le scénario symbiotique : métabolisme et mécanismes moléculaires de la syntrophie H₂
Le scénario le plus soutenu (Figure 5) [7] repose sur une symbiose métabolique (syntrophieCoopération métabolique entre microorganismes reposant sur l’échange de composés tels que H₂, CO₂, formiate ou acétate, dont la consommation par un partenaire permet le métabolisme de l’autre. Ce type d’interaction est considéré comme une étape clé dans les scénarios de l’origine de la cellule eucaryote.) entre une archée Asgard H₂-dépendante et une α-protéobactérie anaérobie facultative [8]. Ce scénario est aujourd’hui largement soutenu par les données disponibles, mais il n’est pas le seul proposé. D’autres modèles diffèrent quant aux mécanismes ayant conduit à l’intégration de la bactérie dans la cellule hôte, tout en reconnaissant le rôle central de l’endosymbiose mitochondriale.
Mécanismes de la syntrophie H₂
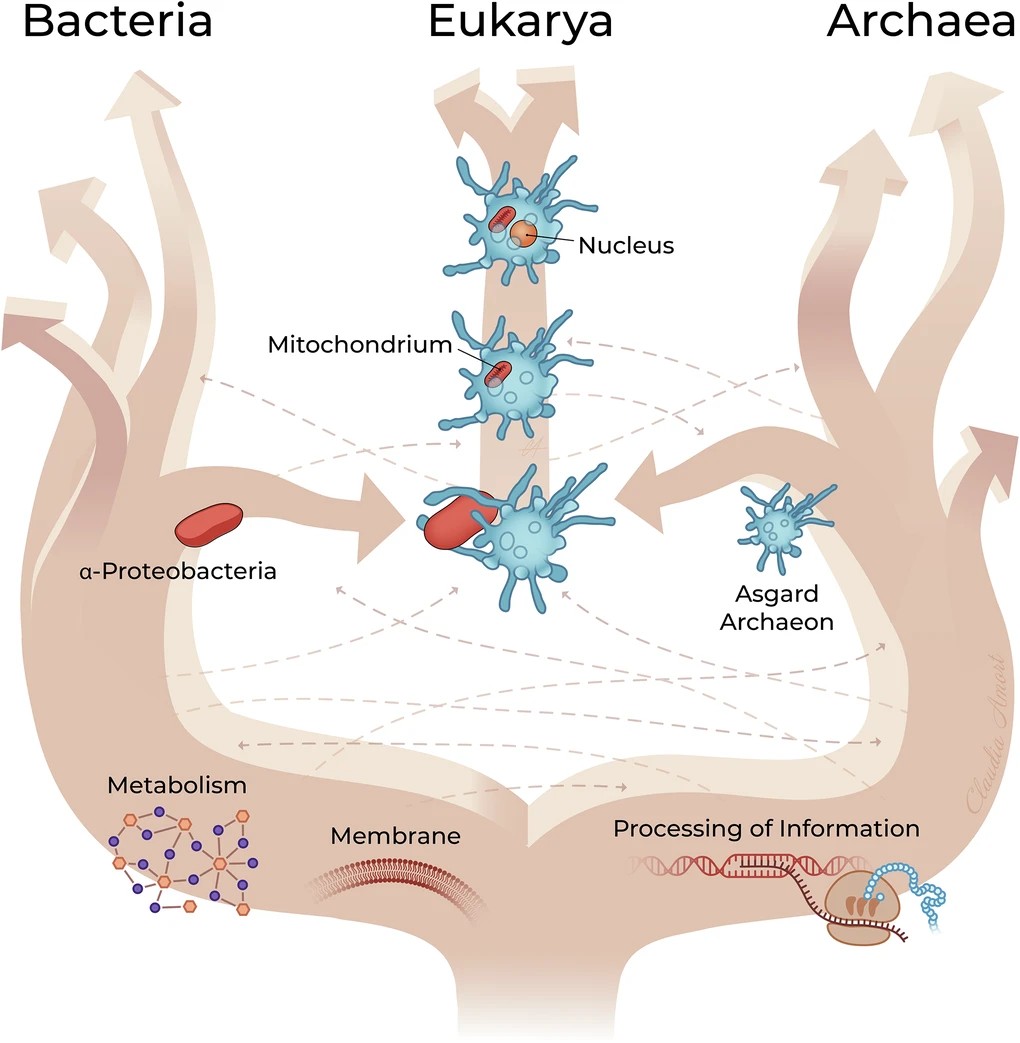
Chez les Asgard, ce métabolisme syntrophique implique des hydrogénases [FeFe] ou [NiFe]. Les deux partenaires pouvaient également échanger directement des électrons grâce à des contacts physiques entre leurs membranes, renforçant leur coopération métabolique. L’archée hôte, utilisant probablement l’acétate ou le méthane, dépendait fortement de cet H₂ exogène. L’α-protéobactérie ancestrale était capable de fermentation productrice d’H₂ en anaérobiose et de respiration en présence d’oxygène. Cette flexibilité a permis une association de plus en plus intime, passant de la syntrophie extracellulaire à l’endosymbiose [8].
L’internalisation a pu suivre le modèle Entangle–Engulf–Endogenize (E³), proposé à partir des observations réalisées chez Candidatus Prometheoarchaeum syntrophicum (voir Figure 5) [11]. Selon ce modèle, les protrusions membranaires de l’archée auraient progressivement entouré le partenaire bactérien avant son internalisation selon un processus inconnu, mais différent de la phagocytose classique (les archées n’ont pas de machinerie phagocytaire connu).
L’intégration de l’α-protéobactérie a profondément transformé la cellule hôte. En devenant la mitochondrie, elle lui a permis de produire beaucoup plus d’énergie que par la fermentation, le mode de fonctionnement des premiers organismes. Les mitochondries fabriquent ainsi environ quinze fois plus d’ATP – la principale molécule énergétique de la cellule – à partir d’une même quantité de glucose. Ce gain d’énergie a favorisé l’augmentation de la taille des cellules et l’apparition de structures complexes, comme le cytosquelette, le système endomembranaire et, plus tard, le noyau.
Contrairement aux chloroplastes, acquis à plusieurs reprises, la mitochondrie ne semble être apparue qu’une seule fois dans l’histoire du vivant : cette acquisition unique constitue l’événement fondateur dont descend l’ensemble de la lignée eucaryote actuelle.
2.3. Les transferts de gènes : un processus massif et continu
L’intégration s’est accompagnée de transferts massifs de gènes de la bactérie vers le génome nucléaire de l’hôte (voir Figure 5). La très grande majorité (plus de 99 % selon les estimations) des gènes de l’ancêtre α-protéobactérien a été perdue ou transférée vers le noyau [12] :
- Relocalisation de fragments d’ADN bactérien dans le noyau ;
- Acquisition de séquences régulatrices eucaryotes et d’un peptide de transit ;
- Perte de la copie redondante du gène de l’organite.
Ces étapes ne se produisent pas simultanément mais au cours d’un long processus évolutif. Ces transferts ont concerné non seulement des gènes métaboliques mais aussi des gènes impliqués dans la réplication, la transcription et la traduction mitochondriales. Ils ont enrichi considérablement le génome nucléaire, contribuant à faire du génome nucléaire une véritable chimère génétique [13], tout en rendant l’organite dépendant du noyau pour la majorité de ses protéines. Ce processus s’est poursuivi sur des centaines de millions d’années et constitue un exemple majeur d’évolution par endosymbiose.
2.4. Le débat sur l’oxygène et la chronologie
Le rôle exact de l’oxygène dans cette transition évolutive reste discuté. Les débats portent moins sur l’existence de la symbiose que sur les conditions environnementales dans lesquelles elle s’est produite et sur le métabolisme des deux partenaires au moment de leur association. Les reconstructions ancestrales indiquent un ancêtre commun anaérobie H₂-dépendant [9]. L’α-protéobactérie ancestrale était elle-même anaérobie facultative. Cependant, des travaux récents montrent que certains Asgard pouvaient tolérer de faibles concentrations d’oxygène. La mitochondrie n’aurait donc pas été uniquement une réponse à l’augmentation globale de l’oxygène atmosphérique, mais un amplificateur énergétique qui a permis d’exploiter efficacement cet oxygène là où il était disponible localement.
Une partie de la complexité eucaryote a pu émerger avant l’acquisition mitochondriale chez l’hôte Asgard, comme le suggèrent la présence de protéines homologues des ESP et certaines caractéristiques cellulaires des archées Asgard actuelles. Le débat sur la chronologie précise reste ouvert.
2.5 Et le noyau ?
La question de l’origine de la cellule eucaryote est aussi liée à celle du noyau. C’est une des questions les plus débattues de l’eucaryogenèse. L’hypothèse autogène propose que l’enveloppe nucléaire soit apparue par invagination progressive de la membrane plasmique de l’hôte archéen (sans intervention d’un partenaire externe), tandis que l’hypothèse symbiogénique (ou virale, selon les variantes) propose qu’elle dérive de la fusion ou de l’internalisation d’un autre organisme — une bactérie, ou dans certaines versions un virus à ADN — dont la membrane aurait donné naissance au noyau. Cette séparation entre noyau et cytoplasme aurait pu constituer une réponse évolutive à la nécessité de dissocier l’épissage des ARN de leur traduction, devenue particulièrement importante après les transferts massifs de gènes vers le noyau. C’est alors la pression sélective qui aurait conduit à la fixation de cette compartimentation [9].
La cellule eucaryote est donc le résultat d’une chimérisation profonde : une archée Asgard fournissant l’architecture informationnelle et une partie du cytosquelette, une α-protéobactérie apportant le métabolisme oxydatif via la mitochondrie grâce à une syntrophieCoopération métabolique entre microorganismes reposant sur l’échange de composés tels que H₂, CO₂, formiate ou acétate, dont la consommation par un partenaire permet le métabolisme de l’autre. Ce type d’interaction est considéré comme une étape clé dans les scénarios de l’origine de la cellule eucaryote. H₂ initiale, et des transferts de gènes massifs fusionnant les deux lignées [8], [9] (Figure 6). Cette association a surmonté les limites énergétiques des procaryotes et permis l’émergence d’une complexité cellulaire sans précédent.
Contrairement à l’origine des mitochondries, largement établie, l’origine du noyau demeure l’une des grandes questions ouvertes de la biologie évolutive. Plusieurs hypothèses sont actuellement étudiées, sans qu’aucune ne fasse encore consensus.
3. L’origine endosymbiotique du chloroplaste
3.1 De la phagocytose à l’endosymbiose
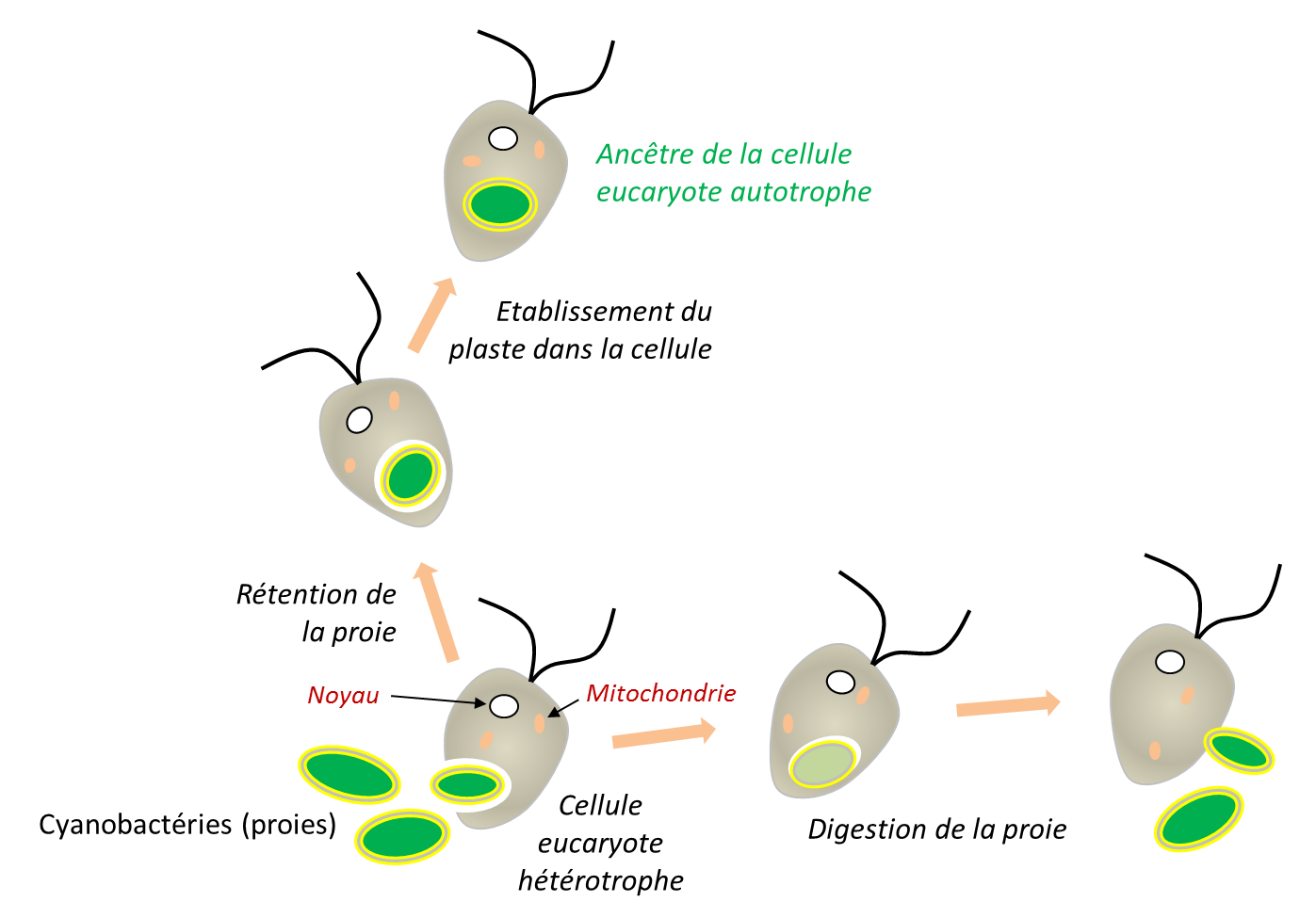
Lors de la phagocytose, la membrane plasmique s’invagine autour de la proie et forme une vésicule d’endocytose, qui fusionne ensuite avec les lysosomes où son contenu est digéré. On a longtemps considéré que la membrane externe des chloroplastes provenait de cette membrane d’endocytose. Les données actuelles suggèrent cependant un scénario plus complexe (Figure 8). Les ancêtres bactériens des chloroplastes (cyanobactéries) et des mitochondries (α-protéobactéries) étaient déjà des bactéries à Gram négatifBactérie caractérisée par une fine couche de peptidoglycane comprise entre une membrane interne et une membrane externe contenant des lipopolysaccharides. Cette organisation est notamment celle des α-protéobactéries, dont est issue la mitochondrie. possédant deux membranes. De plus, la membrane externe des chloroplastes, notamment sa face exposée au cytosol, contient des glycolipides caractéristiques des cyanobactéries [16]. Ces observations suggèrent que la membrane d’endocytose a pu disparaître au cours de l’intégration de l’endosymbiote (Figure 8).
On a longtemps attribué cette remarquable longévité à un transfert de gènes de l’algue vers le génome de la limace. Des séquençages complets du génome d’Elysia chlorotica, plus complets et plus fiables que les premières études, n’ont cependant révélé aucun transfert de ce type. La persistance de ces chloroplastes « volés » demeure donc largement inexpliquée [17]. Cet exemple illustre la manière dont une hypothèse scientifique peut être révisée à la lumière de nouvelles données.
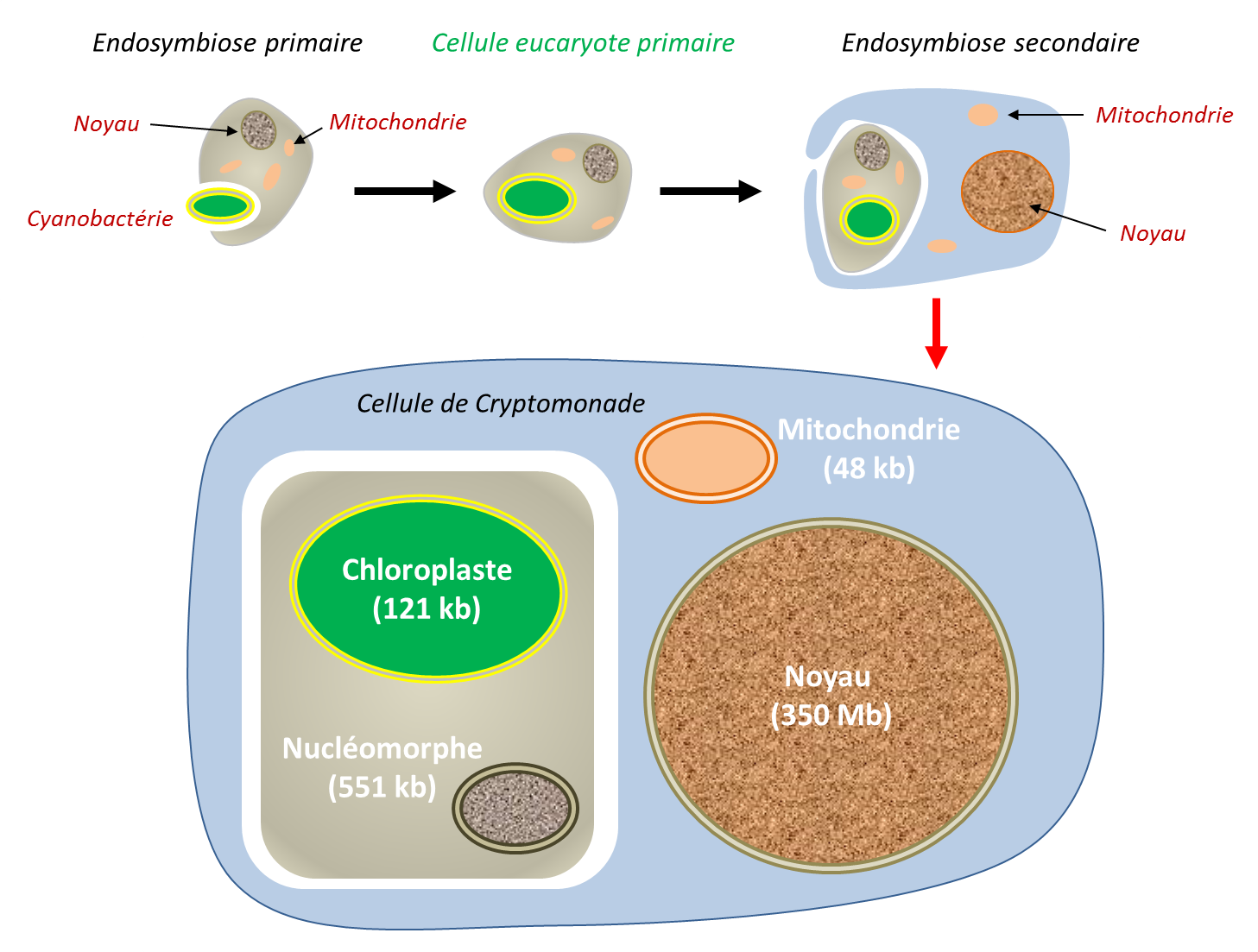
3.2 Endosymbioses primaires, secondaires et tertiaires
L’endosymbiose ne s’est pas produite une seule fois au cours de l’évolution. Plusieurs événements indépendants ont conduit à l’apparition de nouvelles lignées d’organismes. Une endosymbiose primaire correspond à l’internalisation d’un procaryote vivant par une cellule eucaryote. Les chloroplastes des glaucophytes, des algues rouges et de la lignée verte dérivent tous d’une telle endosymbiose impliquant une cyanobactérie. Chez certains eucaryotes adaptés aux milieux anaérobies, les mitochondries ont profondément évolué sans jamais disparaître. Elles ont donné naissance à des organites dérivés, tels que les hydrogenosomesOrganite dérivé de la mitochondrie, présent chez certains eucaryotes adaptés aux milieux dépourvus d’oxygène. Il produit de l’énergie sous forme d’ATP par un métabolisme anaérobie, avec libération d’hydrogène (H₂), illustrant la diversité évolutive des organites issus de l’endosymbiose mitochondriale. On en trouve chez certains ciliés anaérobies, les Trichomonas, et les champignons. réalisant une fermentation productrice de H2 (par exemple chez certains Ciliés) [18], mais aussi des organites de petite taille, seulement impliqués dans des biosynthèses pour la cellule hôte, les mitosomes [18]. Aujourd’hui, ces organites sont regroupés sous le terme de MROEnsemble des organites issus de l’évolution des mitochondries chez des eucaryotes adaptés aux environnements pauvres en oxygène (milieux anaérobies ou microaérobies). Les MRO illustrent la remarquable plasticité évolutive des mitochondries, qui ont conservé ou modifié leurs fonctions selon les contraintes de leur environnement. Selon les espèces, ils assurent différentes fonctions métaboliques, comme la production d’ATP (hydrogénosomes) ou la biosynthèse des centres fer-soufre (mitosomes). (mitochondrion-related organelles), qui recouvre un continuum de formes dérivées des mitochondries. Certains MRO, comme ceux de Blastocystis, présentent des caractéristiques intermédiaires [19].
Les endosymbioses secondaires correspondent à une réitération du processus, lorsqu’une cellule eucaryote contenant déjà un endosymbiote est internalisée par une autre cellule eucaryote (Figure 8). C’est l’origine des plastes entourés de plus de deux membranes présents chez plusieurs lignées d’eucaryotes : internalisation d’une algue verte chez des Euglènes ; internalisation indépendante d’une algue rouge chez les algues brunes. Des endosymbioses tertiaires, moins fréquentes, ont aussi été décrites. Ces événements constituent autant d’endosymbioses fondatrices de nouvelles lignées évolutives [20],[21].
Une seconde endosymbiose primaire, totalement indépendante de celle qui est à l’origine des plastes des glaucophytes, des algues rouges et de la lignée verte, a été découverte chez l’amibe Paulinella chromatophoraAmibe d’eau douce qui possède un organite photosynthétique issu d’une cyanobactérie acquise par endosymbiose primaire il y a environ 100 millions d’années. Elle constitue un exemple unique d’endosymbiose primaire indépendante de celle à l’origine des chloroplastes des plantes et des algues.. Cette amibe a internalisé une cyanobactérie apparentée aux genres Prochlorococcus et Synechococcus, devenue un organite photosynthétique appelé chromatophore.
Beaucoup plus récente (60 à 140 millions d’années) que l’endosymbiose à l’origine des chloroplastes classiques (plus de 1,5 milliard d’années), cette association constitue aujourd’hui un modèle privilégié pour étudier les premières étapes de la transformation d’un endosymbiote en organite [22].
3.3. De l’endosymbiote à l’organite
Quel que soit le scénario considéré, la réussite d’une endosymbiose dépend ensuite de l’intégration progressive de l’endosymbiote au fonctionnement de la cellule hôte. Cette intégration progressive transforme peu à peu l’endosymbiote en un véritable organite. Celui-ci concerne successivement son génome, son fonctionnement et sa transmission au sein de la cellule hôte.
4. L’intégration du procaryote au sein de la cellule eucaryote
4.1. Une réduction massive des génomes
Toutes les endosymbioses décrites précédemment présentent une caractéristique commune : une forte régression du génome de l’endosymbiote. Par comparaison avec les génomes des protéobactéries libres, les mitochondries ont perdu plus de 99 % de leurs gènes. Les plastes de la lignée verte n’en conservent plus qu’une centaine, contre plusieurs milliers chez les cyanobactéries libres. À l’extrême, les hydrogénosomes et les mitosomes ont même perdu leur génome, tandis que certains plastes très régressés, comme ceux de la plante parasite Rafflesia, n’en possèdent plus non plus [23].
Cette régression résulte notamment de la disparition des gènes devenus inutiles dans le milieu intracellulaire. Ainsi, les cyanobactéries possèdent, comme toutes les bactéries Gram négatif, une couche de peptidoglycane située entre leurs deux membranes. Indispensable dans le milieu extérieur, cette structure devient inutile une fois l’endosymbiote intégré dans le cytoplasme de la cellule hôte, où les variations des conditions osmotiques sont beaucoup plus limitées. Les gènes responsables de sa synthèse ont donc été perdus dans les chloroplastes, à l’exception des glaucophytes [24].
4.2. Le transfert des fonctions et de gènes vers le noyau
Malgré la forte réduction de leur génome, les organites conservent un protéome en grande partie comparable à celui de leurs ancêtres bactériens. Cette apparente contradiction s’explique par le fait que de nombreuses protéines sont désormais codées par le génome nucléaire de la cellule hôte. Synthétisées dans le cytosol, elles sont ensuite importées dans l’organite grâce à une séquence d’adressage appelée peptide de transit. Cette relocalisation des fonctions vers le noyau constitue l’une des innovations majeures de l’intégration endosymbiotique.
Les systèmes assurant cet import représentent eux-mêmes une innovation évolutive remarquable. Constitués de protéines d’origine bactérienne et eucaryote, ils reconnaissent les protéines destinées à l’organite, les transportent à travers les membranes, puis les dirigent vers leur compartiment fonctionnel [25].
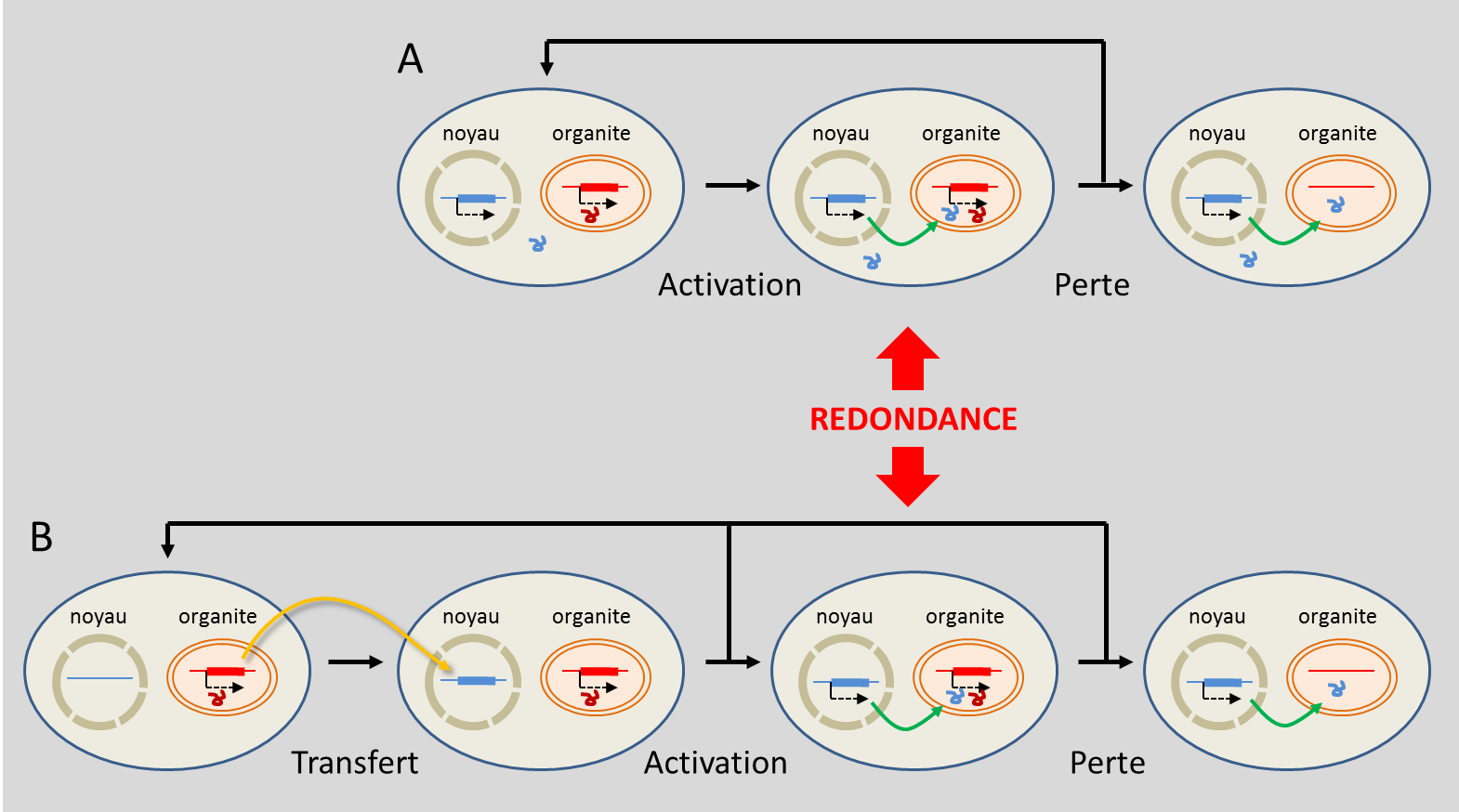
Les transferts d’ADN des organites vers le noyau sont loin d’être exceptionnels. Chez Arabidopsis thaliana, on estime que près de 10 % des gènes nucléaires dérivent, directement ou indirectement, de transferts anciens issus des plastes, souvent suivis de duplications [26]. Leur intégration résulterait vraisemblablement de la dégradation d’organites libérant des fragments d’ADN ensuite incorporés de manière aléatoire dans le génome nucléaire.
La réduction du génome des organites résulte d’un équilibre entre plusieurs forces évolutives. Beaucoup de gènes ont été transférés vers le noyau, tandis que d’autres ont été conservés dans les organites en raison de contraintes fonctionnelles qui font encore l’objet de discussions. Ainsi, l’endosymbiose réduit progressivement les génomes des endosymbiotes tout en enrichissant celui du noyau, qui devient un véritable génome chimérique associant des gènes issus de différentes lignées évolutives.
4.3. La coordination entre l’organite et la cellule hôte
L’intégration d’un endosymbiote ne dépend pas seulement des transferts de gènes. Elle exige aussi une coordination étroite entre la division de l’organite et celle de la cellule hôte, condition indispensable à sa transmission au fil des générations.
Pour être transmis aux cellules filles, les plastes doivent se diviser avant la division de la cellule hôte et être répartis entre les deux cellules issues de cette division. Une multiplication trop rapide leur permettrait de proliférer au détriment de l’hôte ; une division insuffisante conduirait au contraire à leur disparition. L’évolution a donc sélectionné une étroite coordination entre les cycles de division de l’hôte et de l’endosymbiote.
La plupart des protéines impliquées dans la division des chloroplastes dérivent de la machinerie de division des cyanobactéries. Quelques-unes sont toutefois d’origine eucaryote et, fait remarquable, elles sont aujourd’hui toutes codées par le génome nucléaire. Ce contrôle nucléaire constitue l’une des manifestations les plus abouties de l’intégration de l’endosymbiote à la cellule hôte.
L’intégration d’un endosymbiote ne se limite donc ni à son maintien dans la cellule ni au transfert d’une partie de ses gènes. Elle aboutit à une véritable fusion fonctionnelle entre deux partenaires auparavant indépendants. Cette innovation a profondément transformé l’évolution des eucaryotes.
5. La symbiose, moteur de l’évolution ?
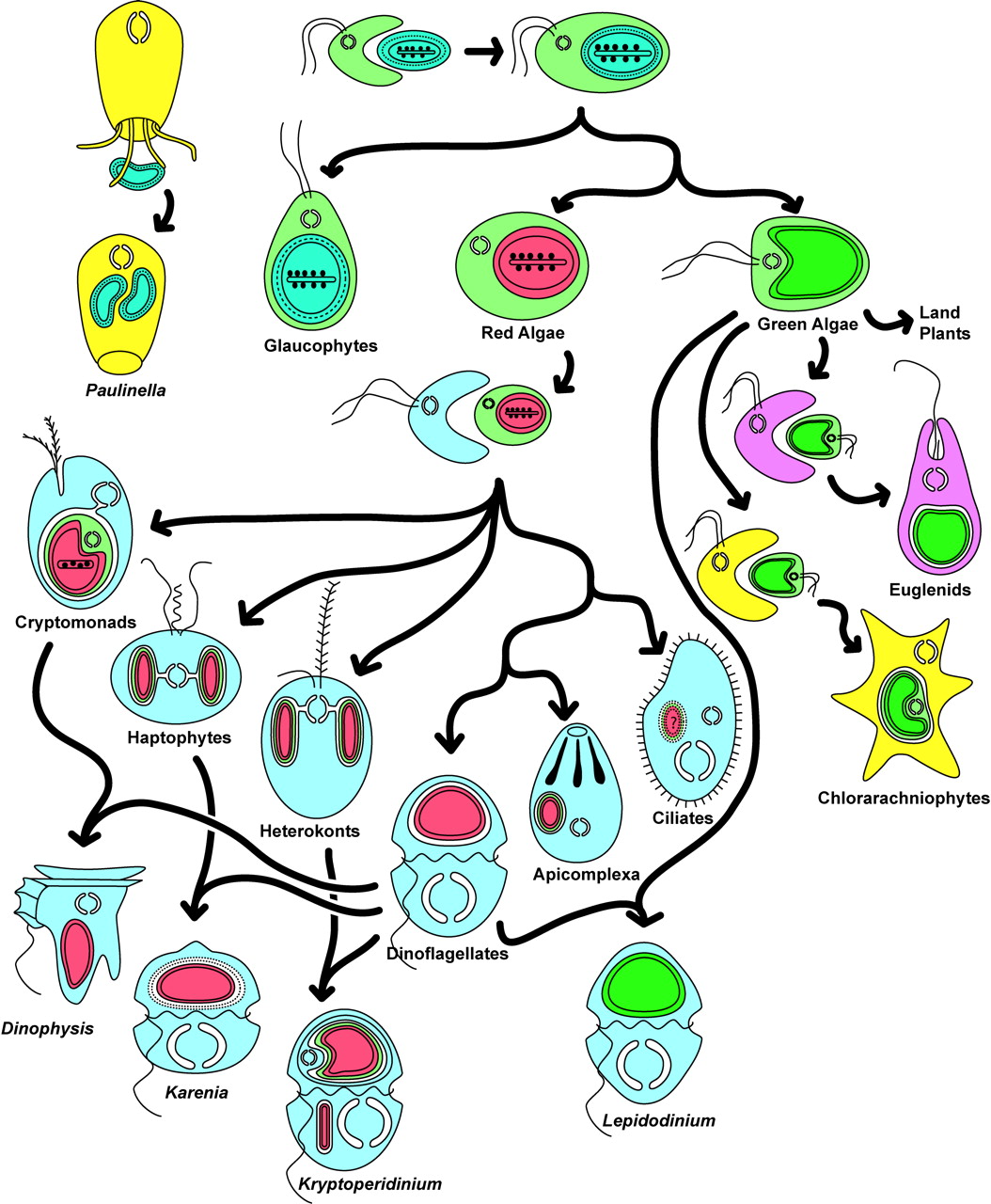
Loin d’appartenir uniquement au passé, ces processus continuent d’être observés. Certaines algues unicellulaires, notamment des cryptophytesOrganismes unicellulaires, photosynthétiques pour la plupart. Leurs chloroplastes sont limités par quatre membranes, signe de l’intégration d’une algue rouge par endosymbiose secondaire. La présence d’un nucléomorphe chez de nombreuses espèces en fait un modèle important pour comprendre l’évolution des organites photosynthétiques complexes. Les Cryptophytes se rencontrent dans de nombreux milieux, en particulier aquatiques (milieux océaniques, eaux douces, eaux interstitielles des milieux terrestres humides). Certaines espèces sont devenues parasites intestinaux de métazoaires. Quelques-unes sont des endosymbiotes de Dinophytes. et des hétérocontesGrande lignée d’eucaryotes regroupant des organismes photosynthétiques (diatomées, algues brunes, chrysophytes…) et non photosynthétiques (oomycètes). Les espèces photosynthétiques ont acquis leurs chloroplastes par endosymbiose secondaire d’une algue rouge, illustrant l’importance des endosymbioses dans l’évolution des eucaryotes., dont les plastes à quatre membranes proviennent d’une endosymbiose secondaire, vivent aujourd’hui comme endosymbiotes dans le cytoplasme de dinoflagellésEucaryotes unicellulaires principalement marins, caractérisés par la présence de deux flagelles. Leur grande diversité métabolique et leurs plastes d’origines variées, acquis au cours de plusieurs événements d’endosymbiose secondaire ou tertiaire, en font un modèle majeur pour l’étude de l’évolution des organites photosynthétiques. ayant perdu leurs propres plastes. Ces associations illustrent la succession de plusieurs événements endosymbiotiques au cours de l’évolution et montrent que l’intégration de nouveaux partenaires cellulaires demeure un processus évolutif en cours.
Les progrès de la phylogénomique, de la microbiologie, de la biologie cellulaire et de la microscopie continuent d’affiner notre compréhension de l’origine des eucaryotes. Si le principe général d’une origine endosymbiotique de la cellule eucaryote est aujourd’hui largement admis, plusieurs questions demeurent ouvertes, notamment l’origine du noyau, la chronologie des événements ayant conduit à LECA et les mécanismes précis d’internalisation de l’α-protéobactérie. Chaque nouvelle archée Asgard découverte, chaque génome séquencé et chaque culture obtenue apportent de nouvelles pièces à ce puzzle évolutif.
Longtemps considérée comme une curiosité biologique, l’endosymbiose est aujourd’hui reconnue comme l’un des principaux moteurs de l’innovation évolutive. En réunissant durablement des organismes auparavant indépendants, elle a permis l’émergence de cellules plus complexes, favorisé l’apparition de la multicellularité et ouvert la voie à la diversification des plantes, des champignons et des animaux. L’histoire de la cellule eucaryote illustre ainsi l’une des plus grandes transitions de l’évolution : celle où la coopération entre organismes est devenue une source majeure de nouveauté biologique.
Références & notes
Image de couverture : [Source Photo © Jacques Joyard]
[1] La figure 2 est créée à partir des données des références suivantes : Lang T. et al. (2000) Autophagy and the cvt pathway both depend on AUT9. J Bacteriol 182, 2125-2133. doi: 10.1128/JB.182.8.2125-2133.2000 ; Spang, A., Saw, J. H., Jørgensen, S. L., Zaremba-Niedzwiedzka, K., et al. (2015). Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature, 521(7551), 173–179. https://doi.org/10.1038/nature14447 ; Zaremba-Niedzwiedzka, K., Caceres, E. F., Saw, J. H., Bäckström, D., Juzokaite, L., Vancaester, E., … Ettema, T. J. G. (2017). Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature, 541(7637), 353–358. https://doi.org/10.1038/nature21031 ; Imachi, H., Nobu, M. K., Nakahara, N., Morono, Y., Ogawara, M., Takaki, Y., … Takai, K. (2020). Isolation of an archaeon at the prokaryote–eukaryote interface. Nature, 577(7791), 519–525. https://doi.org/10.1038/s41586-019-1916-6
[2] Selosse M.A. (2012). Gloire et disgrâce de la théorie endosymbiotique. La Recherche 468: 92-94.
[3] Zaremba-Niedzwiedzka, K. et al. (2017). Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature, 541(7637), 353-358. https://doi.org/10.1038/nature21031
[4] Tobiasson, V., Luo, J., Wolf, Y.I. et al. (2026). Dominant contribution of Asgard archaea to eukaryogenesis. Nature 650, 141–149.
[5] Zhang, J., Feng, X., Li, M. et al. (2025). Deep origin of eukaryotes outside Heimdallarchaeia within Asgardarchaeota. Nature 642, 990–998 https://doi.org/10.1038/s41586-025-08955-7
[6] Richards T.A., Eme L., Archibald J.M., Leonard G., Coelho S.M., de Mendoza A. et al. (2024) Reconstructing the last common ancestor of all eukaryotes. PLoS Biology 22(11): e3002917. https://doi.org/10.1371/journal.pbio.3002917
[7] Schleper, C. & Rodrigues-Oliveira, T. (2026) Asgard archaea: have we found our microbial ancestors? EMBO J 45, 1836–1851.
[8] Martin W.F., Garg S. & Zimorski V. (2015) Endosymbiotic theories for eukaryote origin. Phil. Trans. R. Soc. B370, 20140330. doi: 10.1098/rstb.2014.0330.
[9] Martin W. & Müller M. 1998 The hydrogen hypothesis for the first eukaryote. Nature 392, 37–41.
[10] López-García P & Moreira D. (2019) Eukaryogenesis, a syntrophy affair. Nat Microbiol. 4(7):1068-1070. doi: 10.1038/s41564-019-0495-5.
[11] Imachi, H., Nobu, M.K., Nakahara, N. et al. (2020) Isolation of an archaeon at the prokaryote–eukaryote interface. Nature 577, 519–525. https://doi.org/10.1038/s41586-019-1916-6
[12] Selosse M.A., Albert B. & Godelle B. (2001) Small is successful: selection for reducing organelle’s genome size favours gene transfer to the nucleus. Trends Ecol Evol 16, 135-141.
[13] Tobiasson, V., Luo, J., Wolf, Y.I. et al. (2026) Dominant contribution of Asgard archaea to eukaryogenesis. Nature 650, 141–149. https://doi.org/10.1038/s41586-025-09960-6.
[14] Archibald JM & Keeling PJ (2002) Recycled plastids: a ‘green movement’ in eukaryotic evolution. Trends Genetics 18, 577-584.
[15] Keeling PJ (2004) Diversity and evolutionary history of plastids and their hosts. Am. J. Bot. 91, 1481-1493.
[16] Douce R., Block M.A., Dorne A.J., Joyard J. (1984) The plastid envelope membranes: their structure, composition, and role in chloroplast biogenesis. Subcell. Biochem.10, 1-84, Springer US (Ed.)
[17] Cruz S, Cartaxana P (2022) Kleptoplasty: Getting away with stolen chloroplasts. PLoS Biol 20(11): e3001857. https://doi.org/10.1371/journal.pbio.3001857
[18] Selosse MA & Loiseaux-de Goër S (1997) La Saga de l’endosymbiose, La Recherche 296, 36
[19] Stairs C.W., Leger M.M., Roger A.J. (2015) Diversity and origins of anaerobic metabolism in mitochondria and related organelles. Philos. Trans. R. Soc. B 370, 20140326 ; voir aussi Stechmann A. et al. (2008) Organelles in Blastocystis that blur the distinction between mitochondria and hydrogenosomes. Curr. Biol. 18, 580-585.
[20] Lefèvre T., Renaud F., Selosse M.-A., Thomas F. (2010). Evolution des interactions entre espèces, in F. Thomas, T. Lefèvre & M. Raymond (ed.), Biologie évolutive, p. 530-613. De Boeck, Paris.
[21] Keeling PJ (2010) The endosymbiotic origin, diversification and fate of plastids. Phil. Trans. R. Soc. B 365, 729-748
[22] Marin B., Nowack E.C.M., Melkonian M. (2005) A plastid in the making: evidence for a second primary endosymbiosis. Protist 156, 425-432 ; voir aussi Delaye L., Valadez-Cano C., Pérez-Zamorano B. (2016) How really ancient is Paulinella chromatophora? PLoS Curr. Tree of Life 8.
[23] Molina J. Hazzouri K.M., Nickrent D. et al. (2014), Gisela P. Concepcion, Michael D. Purugganan, Possible Loss of the Chloroplast Genome in the Parasitic Flowering Plant Rafflesia lagascae (Rafflesiaceae), Molecular Biology and Evolution, 31,4, Pages 793–803, https://doi.org/10.1093/molbev/msu051
[24] Glaucophytes : Organismes unicellulaires présentant des flagelles (2 de de longueur inégale) vivant dans les mares d’eau douce des régions tempérées.
[25] Selosse MA, Albert B et Godelle B (2001) Small is successful : selection for reducing organelle’s genome size favours gene transfer to the nucleus. Trends Ecol Evol 16, 135-141.
[26] Jarvis P (2004) Organellar Proteomics: Chloroplasts in the Spotlight. Current Biology 14, R317-9. http://www.cell.com/current-biology/references/S0960-9822%2804%2900231-3
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Pour citer cet article : SELOSSE Marc-André, JOYARD Jacques (20 juillet 2026), Symbiose et évolution : à l’origine de la cellule eucaryote, Encyclopédie de l’Environnement. Consulté le 3 août 2026 [en ligne ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/vivant/symbiose-evolution-lorigine-de-cellule-eucaryote/.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.


