




温度对光合作用的影响

陆生植物经常受到温度剧烈变化的影响,无论是在极地还是在炎热的沙漠地区,温度的变化幅度可以达到40 °C甚至更高。陆生植物扎根后就几乎不能移动,因此,它们必须应对原位环境的变化。植物通过光合作用同化二氧化碳,这是碳进入生物圈的起点。光合作用正常工作的温度范围是多少?光合作用对快速和缓慢的温度变化如何响应?不同植物的响应有何差异?限制光合作用的生理过程是什么?这些关键问题都需要我们在全球变暖的背景下加以考虑。
1. 植物生产与气候变化
从目前温室气体排放增加的趋势来看,未来50年内大气温度可能升高2-3°C(见“被人类活动干扰的碳循环”)。与此同时,热浪和极端高温事件将更加频发,持续时间也会更长[1]。农业生产和森林的功能将受到巨大的影响。基于大规模观察的模型表明,在缺乏农业适应性措施的情况下,生长季温度每升高1°C,作物产量下降可达17%[2]。
高等植物的生产力尤其(但不只是[3])依赖叶片的光合作用(参见“揭示光合作用的真相”以及“光合作用的碳代谢路径”)。二氧化碳(CO2)进入叶片后在叶绿体中被还原,同时伴随着氧气(O2)的产生。二氧化碳几乎完全是通过保卫细胞进入叶片的(图1),每一个二氧化碳分子进入叶片,就会蒸发掉50-300个水分子,具体数值取决于植物种类和状态。蒸腾损失的水分除了其他功能外,还能使叶片冷却(参见“叶片蒸腾和热保护”)。
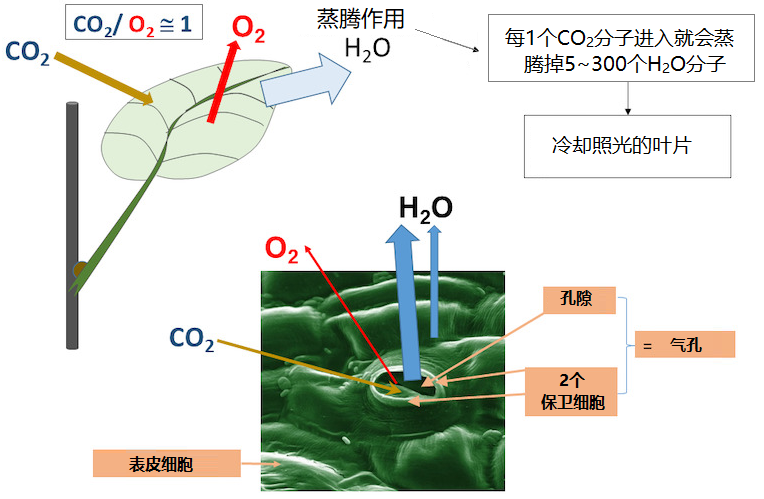
叶片将太阳能转化为化学能,与任何能量转化器一样,它需要一个持久工作的冷却系统。
当前,气候变化正在发生,了解温度对光合作用的影响则更为必要。
2. 光合作用的最适温度
2.1. 温度响应图
光合作用中二氧化碳的吸收速率随温度而变化。大多数情况下,当温度在10-34 °C之间变化时,光合速率也会发生快速可逆的变化,并出现一个最大速率:即光合作用的最适温度。
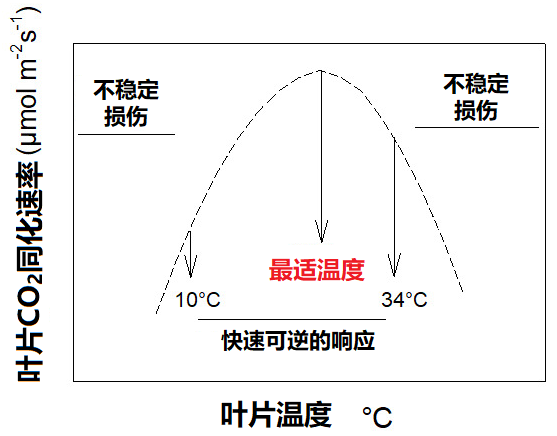
10°C以下和34°C以上对植物来说属于胁迫条件,它们会启动保护机制,以应对此类极端温度。在此胁迫条件下,叶片的二氧化碳同化过程往往是不稳定的,甚至可能停止(图2)。
2.2. 基于环境平均温度的最适温度
在寒冷环境或寒冷季节生长的植物,低温下的光合速率更高;相反,在温暖环境或温暖季节生长的植物,高温下的光合速率更高。

例如,南极仅有两种开花植物南极发草(Deschampsia antarctica,图3)和南极漆姑草(Colobanthus quitensis),它们的二氧化碳同化的最适温度都在8~15°C之间[4],而来自中美洲炎热沙漠的植物(Tridestomia oblongifolia)的最适温度在45°C左右,这可能是有花植物的世界纪录。
2.3. 对环境温度条件的适应
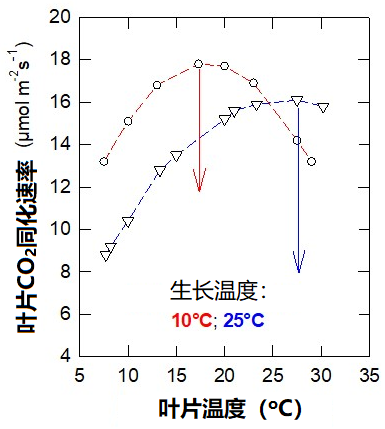
生长在不同温度下的同种植物个体,其光合作用对温度的响应也存在差异。图4显示了在10 °C和25 °C下生长的豌豆的二氧化碳吸收速率。
第一种情形为在10 °C下培养植物,其叶片光合作用的最适温度约为16 °C;而在第二种情形下,植物的培养温度是25 °C,此时的最适温度则高于25 °C。而且在较低温度下,在10 °C条件下生长的植物二氧化碳吸收速率更高。
在这种情况下,适应较冷的温度条件对植物来说是有利的。
2.4. 可以快速适应

例如,中东沙漠(内盖夫的瓦迪拉姆)中的一种灌木雷姆斯(Hamada scoparia)的光合作用随温度的季节变化而变化:最适温度从早春的29 °C升高到夏季的41 °C,再回落到秋季的28 °C。

光合最适温度的变化甚至可以更快,幅度也更大。例如,在加利福尼亚海滨的加州脆菊木(Encelia california)株丛[5],当其生长温度从30 °C(昼夜恒定)变为白天15 °C、夜间2 °C,只需3天,其光合最适温度就会降低约10 °C。
一般来说,无论是还在生长的叶片还是已经成熟的叶片,都可以测量到这样的变化,生长中的叶片响应幅度往往更大。
2.5. 热敏植物与冷敏植物的比较
生活在冷凉环境的植物物种(或生态型[6])对高温的适应会伴随着最适温度的升高,同时光合速率往往会降低。
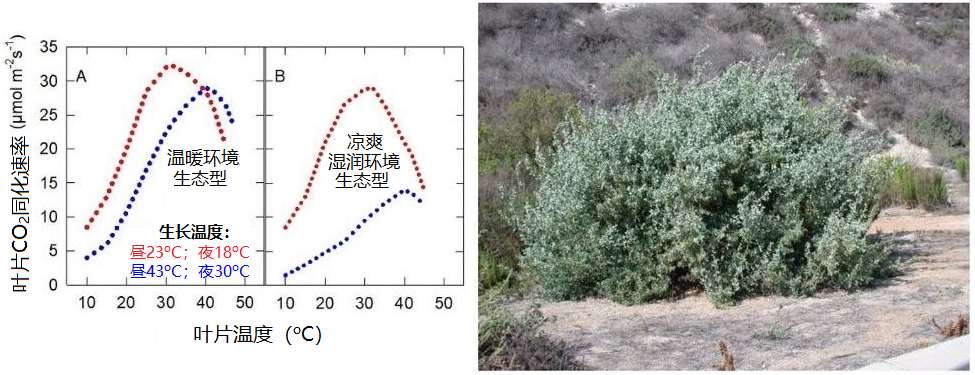
例如,裂叶滨藜(Atriplex sabulosa)就是如此。这种变化的意义值得思考。而对于那些严格适应温暖环境的植物来说,情况可能正好相反,如Tridestomia oblongifolia。图7展示了大滨藜(Triplex lentiformis)的情况,它是一种多年生密丛生小灌木[7],既分布在加州的死亡谷,也生长在凉爽湿润的沿海地带:
- 当生长温度为白天23 °C、夜间18 °C(图7中红色部分) 时,沙漠生态型(图7A)和滨海生态型(图7B)的二氧化碳吸收对温度变化的反应几乎相同。
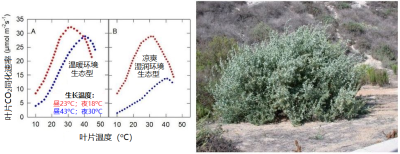
图7. 左图为生活在温暖环境(A)和凉爽湿润环境(B)的两种大滨藜(Atriplex lentiformis)生态型二氧化碳吸收速率的差异。[图片来源:作者自制,见Pearcy(1977)]。右图为大滨藜(盐木)[图片来源:Forest & Kim Starr, CC BY 3.0, 通过维基共享资源]
2.6. C3植物和C4植物的比较

C3植物是最早出现的植物类群,约占现存植物物种的85%。它们主要生长在凉爽湿润的环境(或季节)。例如,树木中除了极个别外,其他都是C3植物(阅读“光合作用的碳代谢路径”)(图8)。
C4植物始现于第三纪末期,仅占现存植物物种总数的5%。它们更喜爱炎热、干燥的环境(或季节)(参见“恢复热带稀树草原和热带草本生态系统”)。玉米和甘蔗就是C4植物。
平均而言,C4植物的最适温度高于C3植物。
但是,C3植物具有极强的适应性。事实上,它们的最适温度范围很宽,从大约7 °C到35 °C左右,而C4植物的最适温度一般在30-40°C之间,只有很少例外。而且当温度低于20 °C时,C4植物的平均光合速率低于C3植物。
3. 二氧化碳同化是温度响应特征不同的诸过程相互作用的结果
捕集天线对光的吸收(图9)和将光能转移到PSII反应中心的过程对温度不敏感。
温度敏感的过程包括:
- 二氧化碳从环境空气到叶绿体的扩散过程:扩散速度随着温度的增加而提高。
- 1,5-二磷酸核酮糖(RuBP)固定二氧化碳的过程,RuBP是一种由5个碳原子构成的糖(“破译本森-巴萨姆-卡尔文循环”)。
- 从PSII到PSI的电子传递过程。

RuBP的再生是通过本森-卡尔文循环(Benson-Calvin cycle)(这是光合作用的“生物化学”过程)完成的,该循环需要电子传递生成的还原力(以NADPH的形式),还需要ATP,ATP是类囊体腔内积累的质子跨膜进入叶绿体基质时,推动ATP酶合成的(图9)。
生成还原力、合成ATP以及电子传递三个过程的温度敏感性相当。
4. C3和C4植物中,决定CO2同化最适温度的过程是什么?
4.1. 与光系统活性及此产生的电子传递过程无关
当存在人工电子受体时,对离体类囊体测量(见图9中的图例)发现,电子传递随温度升高而加快,明显存在最适温度,大约为30 °C,与二氧化碳饱和时叶片吸收CO2的最适温度相当[9]。PSII活性的最适温度与电子转移链的最适温度相同。
- PSI活性不受高温抑制(30°C以上,45°C以下)。高温时,其活性保持稳定甚至有所增加:是PSII的活性限制了电子传递链的活性。
- PSII对高温非常敏感,高温会破坏PSII中使水氧化的蛋白质复合物(见图9)。
C3和C4植物中电子传递对温度的反应相似。但是,这两类植物的光合器官在结构上存在着差异(参见:“光合作用的碳代谢路径”)。
因此,能量供应并不能解释这两类植物光合最适温度的差异,对能量的使用方式不同才是决定性因素。
4.2. 这是答案吗?大气中的氧气对C3和C4植物二氧化碳同化的影响
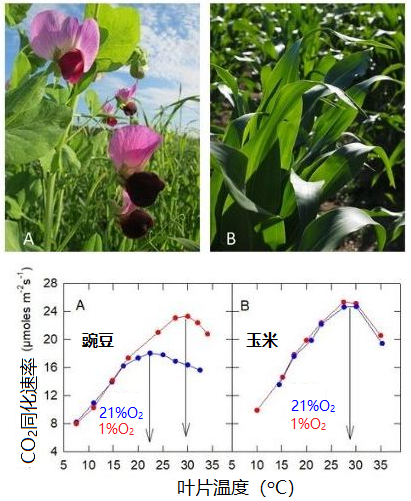
- 在含有21%的O2(+ N2)+ 360 μL/L CO2的正常空气中[10]:玉米(C4植物)的最适温度为27 °C,而豌豆(C3植物)仅为22 °C(图10),C4植物的最适温度高于C3植物(另见2.6节)。
- 在含有1% 的O2(+ N2)+ 360 μL/L CO2的低氧空气中:玉米的二氧化碳吸收不受影响,而豌豆的二氧化碳吸收速率在17°C以上时得以提高,并且最适温度提高到与玉米接近。
- 当叶温很高时,大气中的氧会抑制C3植物的二氧化碳吸收,但对C4植物没有影响(或影响可以忽略)。
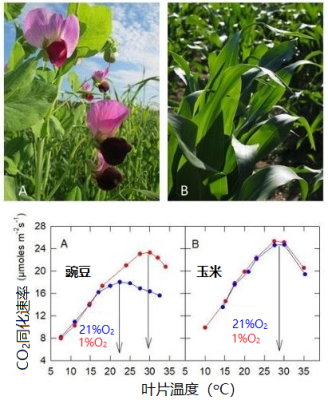
图10. 豌豆(A, Pisum sativum)和玉米(B, Zea mays)叶片二氧化碳吸收速率与叶温的关系。植物生长于20±2 °C的自然光下。[图片来源:作者自制图表和 Pixabay免版税图片]
4.3. Rubisco的性质是差异的关键
- C3植物的情形
二氧化碳和氧气竞争Rubisco的活性位点:该酶同时具有羧化酶功能和加氧酶功能。二氧化碳通过其羧化酶功能进入本森-卡尔文循环的同时,其氧化酶功能导致光合氧气固定,这是光呼吸代谢途径的起点(图11;另见:“光合作用的碳代谢路径”)。
当环境空气中氧气含量较低(例如1%)或二氧化碳含量较高时,二氧化碳会占据大量的Rubisco活性位点。
如果氧气含量增加,或者二氧化碳含量减少(释放出Rubisco酶的活性位点,并被氧气占据),该酶就会主要固定氧气。
在正常的空气中,当温度升高时氧气的固定增加(因而二氧化碳的固定减少),原因有两个[11]。
一是Rubisco对二氧化碳的亲和力下降幅度大于对氧气的亲和力,因而有利于氧气吸收。
二是二氧化碳在水中的溶解系数比氧气下降更多,导致叶绿体中二氧化碳的含量下降比氧气快,这也有利于氧气固定。
在缺氧的大气中(图10),氧气和二氧化碳对Rubisco活性位点的竞争会大大减弱,捕集的光能主要用于二氧化碳的吸收,二氧化碳吸收速率会随着叶片温度的升高而增加,直到30 °C左右达到最大,然后随着能量的减少而降低(见第4.1节)。
当温度较低时,在正常空气中的氧气对光合二氧化碳固定的影响(图11)非常低(甚至为零):此时二氧化碳在羧化位点上的竞争更有优势。
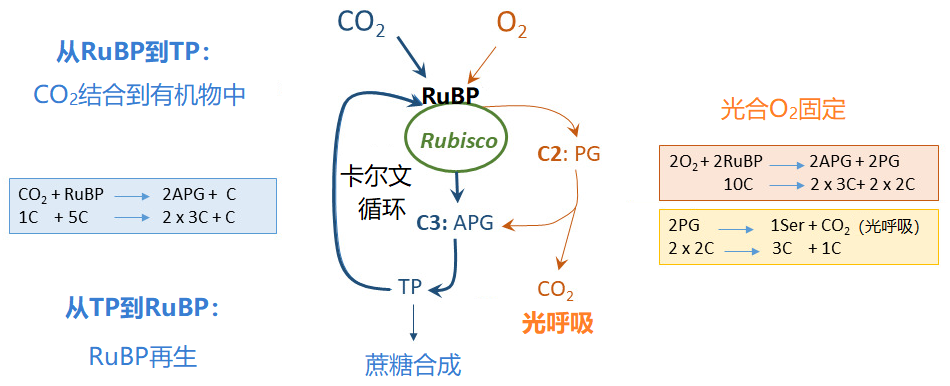
另一方面,当温度升高时,这些活性位点上的竞争有利于氧气的固定,光系统捕获的太阳能越来越多地被氧气固定所消耗,而非用于二氧化碳的固定。二氧化碳固定速率在22 °C左右达到最大值。
- C4植物的情形
二氧化碳通过一种对氧不敏感的机制在Rubisco附近富集,在不同C4植物叶片中含量可以达到800到2000 μL/L:也就是比当前大气中的含量高2到5倍。
在此条件下,由于Rubisco的活性位点全部被二氧化碳占据,光合作用对氧气的固定作用很弱,甚至完全没有。因此,即使叶片温度升高,光系统捕集的太阳光能也仅用于二氧化碳固定,这就是C4植物的光合最适温度较高的原因。
C4植物是在第三纪末期全球大气二氧化碳浓度下降的过程中,从C3植物演化而来的[12]。
大气CO2浓度的降低使得C3植物Rubisco的加氧酶功能得以“发挥”,并通过光呼吸途径损失固定的碳。
因而,建立二氧化碳富集机制就成了一个优势,可以防止光呼吸碳损失。我们现在还能够发现C3和C4“过渡类型”的植物物种。
5. C3光合途径的最适温度受某些环境因子的调控
5.1. 大气二氧化碳浓度
光合最适温度随环境二氧化碳含量的增加而提高。如图12所示,从二氧化碳含量为100 μL/L时的10 °C左右提高到含量为800 μL/L时的30 °C以上。
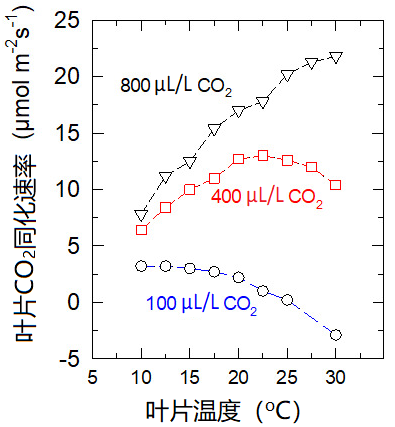
这一效应可以用二氧化碳和氧气竞争Rubisco的活性位点来解释:在二氧化碳浓度为800 μL/L时,活性位点主要被二氧化碳占据;而在二氧化碳浓度为100 μL/L时,氧气会占据大多数的活性位点。

在大气二氧化碳浓度稳步增加的情况下(图13),C3植物的最适温度有望提升,但这并不意味着植物生产力的提高(参见第1部分注释3):因为像干旱一样,高温时期肯定会更频繁地出现。
5.2. 缺水
光合工具叶绿体本身有较强的抗旱性,叶绿体可以保持Rubisco吸收二氧化碳和固定光能的能力,直到叶片失去其含水量的30%[13]。
- 在这种缺水情况下,气孔逐渐关闭(参见“叶片蒸腾和热保护”),二氧化碳吸收会减少。气孔关闭减缓了二氧化碳进入叶片的速度,导致叶肉细胞中二氧化碳含量减少。
- 然而,叶绿体中的氧气浓度仍然很高。事实上,与二氧化碳(约 400 μL/L)相比,大气中氧气的浓度(21%或210 000 μL/L)非常高。在任何情况下,即使气孔关闭,也会有相当数量的氧气通过表皮进入叶片内部。
- 因此,二氧化碳和氧气之间对Rubisco活性位点的竞争对氧气有利。
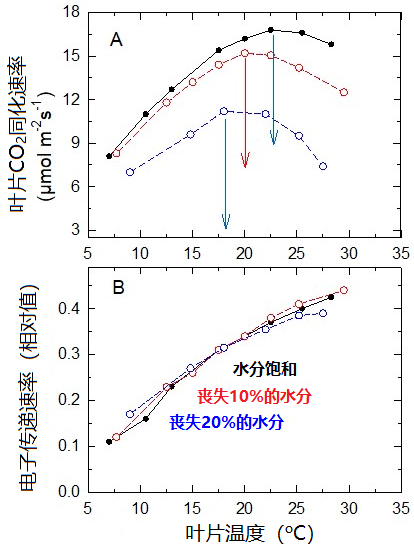
因此,C3植物的叶片在逐渐失水的过程中,其光合作用的最适温度也必须降低。
如图14A所示,豌豆叶片的最适温度从叶片水分饱和时的23 °C,下降到失去20%水分时的17 °C。
在实验中的水分丧失范围内(图14B),类囊体膜上的电子传递速率不受影响。当水分损失达20%时,光系统捕集的太阳能主要用于氧与RuBP结合[14],导致光呼吸增加。
6. 为什么无论温度较最适宜温度升或降,二氧化碳吸收速率总是降低的?
6.1. 当温度低于最适温度时
可能有几个原因不同程度地导致了同化速率降低:
- RuBP周转速度降低。控制RuBP周转的一些酶的活性降低,尤其是果糖1,6-二磷酸酶(见图9和11)。
- 叶绿体中的磷被磷酸化化合物固定。由于蔗糖合成受到抑制,磷酸丙糖输出终止(或减少),使得无机磷酸盐不再与磷酸丙糖交换进入叶绿体,导致ATP合成因无机磷缺乏而下降。
- 抑制电子传递链(见第4.1节),导致能量 (还原力和ATP) 产生减少。
在C4植物中,光合作用对低温的响应主要受Rubisco活性的影响,当然,参与二氧化碳固定的其他酶对低温敏感也是众所周知的。
6.2. 当温度高于最适温度时
在C3植物中,随着温度升高,光呼吸增加, PSII产生的电子中用于同化二氧化碳的比例下降。除此之外,还有其他因素也在起作用:(1)即使在很少或没有光呼吸的大气条件(环境氧气含量为1%)下测量,也会发现光合速率随温度升高而降低;(2)在正常的大气条件下C4植物也有相同的表现 (图10)。可能的原因有:
- PSII活性的降低导致了从PSII到PSI的电子传递速率减慢。
- Rubisco要发挥作用,必须被一种叫做Rubisco激酶的酶激活。当温度超过约33°C时,该酶的活性就会下降(不过,在一些生长期会经历高温的植物中存在耐高温的激酶[15])。但是,由于激酶本身的激活必须依赖电子传递的过程,因此不能排除电子传递也参与限制Rubisco活性[15]。
- 随着温度的升高,与RuBP结构相近的Rubisco酶抑制剂(木糖糖-1,4-二磷酸)的合成量增加,Rubisco的“催化失效”越来越明显(见图9和11)。
在C4植物 (如玉米)中,负责二氧化碳富集在Rubisco周围的酶的激活及其活性对高温不太敏感。因此,C4植物在最适温度以上时,二氧化碳同化速率随着温度升高而下降。
7. 植物的低温(≤10 °C左右)和高温(≥37°C左右)锻炼
当植物停留在低温或高温环境下时,除了引发光合作用的上述变化外,还能增强它们对致死温度(霜冻和高温)的抵抗力,这就是抗性锻炼。
在此过程中,温度和光照共同作用,引起代谢变化,这个变化有时非常快(从几分钟到几小时)。
因此,通过改变光周期的长度或光谱中的红光成分[16],可以在正常温度下实现抗寒锻炼。然而,要使抗性能力完全表现出来,仍然需要低温处理。同样地,低温并缺乏光照的条件也会不同程度地降低抗寒锻炼的效果。
- 高温:产生信号激活分子伴侣性蛋白(热激蛋白,HSPs)的合成,修复变性蛋白,还会防止其凝固,甚至做上标记,使其降解。
- 低温:也会激活分子伴侣性蛋白的合成,随后(i)合成干扰冰晶形成的“抗冻”蛋白和(ii)增加糖的合成,以提高细胞内的渗透压。
请注意,抗性锻炼过程中的信号通路以及它们如何相互作用以诱导基因组反应,目前已知内容有限。读者可以阅读“了解更多”中列出的参考资料和本文的推荐文献,进一步探索这一不断发展的主题。
8. 温度对光合作用的影响:总结图
总结图(图15)根据温度变化的速度和程度,将温度对光合作用的影响进行了分类。请注意,抗性锻炼可以使植物的多年生叶片得以在极端温度条件下维持活性,因而最大限度地减少光能损失。

当前气候变化速度如此之快,我们必须更深入地研究植物对其环境的反应:以期维持足够的初级生产力,保持生物圈功能的可持续性。
9. 需要记住的信息
- 叶片对二氧化碳吸收有最适温度,与接近其生长环境的平均温度。
- 当环境条件被持久地改变时,最适温度也会迅速改变:这是一个驯化过程。
- C3植物的最适温度总体上低于C4植物:这主要是由于C3植物的Rubisco在光合固碳的同时还固定大气中的氧气。
- C3植物的最适温度受环境空气中的二氧化碳浓度影响:当二氧化碳浓度高时,它们的最适温度与C4植物相当。
- 叶片光合最适温度受叶片水分状况影响。
- 植物经受低温或高温条件时,会通过生理过程的抗性锻炼应对致命温度。这些过程涉及蛋白质合成,以及叶绿体和细胞膜流动性的变化。
参考资料及说明
封面图片:亚利桑那州索诺兰沙漠的日落。[来源:免版税/ Pixabay]
[1] Meehl GA、Stocker TF、Collins WD、Riedlingstein P、Gaye AT、Gregory JM、Kitoh A、Knutti R、Murphy JM、Noda A & Raper SCB(2007)《气候变化2007:物理科学基础》。第一工作组对政府间气候变化专门委员会第四次评估报告的贡献。剑桥大学出版社
[2] Yamori W, Hikosaka K & Way DA(2014)。C3、C4和CAM植物光合作用的温度响应:温度驯化和温度适应。光合作用 119, 101- 117。
[3] 例如,当植物遭受干旱时,叶片生长首先被抑制,导致碳固定速率下降。但是叶绿体中的二氧化碳固定过程没有受到抑制。Boyer JS (1970) Plant Physiol.46, 233-235
[4] 这里给出的最适温度是在含21%的氧气和约400 μL/L二氧化碳的“正常空气”中测量得出的。其他条件则标出了氧气和二氧化碳的含量。当接近饱和光强、含21% 氧气的空气条件下,光合速率的二氧化碳饱和点约为1200 μL/L。很多时候在测量过程中空气蒸发能力也会影响光合速率,这个参数是通过叶片周围空气中水汽分压的饱和亏缺来估算的。
[5] 通过营养繁殖从同一个体长出的不同植株,具有相同的基因。
[6] 生态型:生长在不同环境的同种植物,即使种植在相同的条件下,它们在从种子萌发到开花结实的过程中,也会表现出不同的生理特征。
[7] 它可以吸收土壤深处的地下水,因此被称为地下水湿生植物。
[8] Pearcy RW(1971)。triplex lentiJormis (Torr.)光合和呼吸二氧化碳交换对生长温度的适应寺庙。植物生理。59,795-799
[9] Yamasaki T, Yamakawa T, Yamane Y, koike H, Satoh K & Katoh S.(2002)冬小麦光合作用的温度适应及其光系统II电子传递的相关变化。Plant Physiol. 128 1087-1097
[10] 见2.2节的注释#4
[11] Jordan DB & Ogren WL(1984)。的有限公司2/ 氧气 核酮糖1,5-二磷酸羧化酶/加氧酶的特异性。二磷酸核酮糖浓度、pH和温度的依赖性。161年足底,308 – 313
[12] Ehleringer JR, Sage RF, Flanagan LB & Pearcy RW(1991)。气候变化与C4光合作用的演化。生态与进化趋势6,95-99
[13] Cornic G & Massacci A(1996)。干旱胁迫下的叶片光合作用。In Advances In光合作用(第5卷)光合作用与环境,347-366。尼尔R·贝克(主编)Kluwer学术出版社Dordrecht。
[14] Cornic G, Badeck F-W, Ghashghaie J & Manuel N(1999)。温度对CO净吸收二氧化碳、气孔导度的影响2 以及脱水豌豆叶片光系统II光化学的量子产率。In Sanchez Dias M, Irigoyen JJ, agureolea J & Pithan K (eds)欧洲凉爽潮湿地区的作物发育。欧洲共同体。ISBN 92-828-6947-4。
[15] Crafts-Brandner SJ, van de Loo FJ & Salvucci ME(1997)。两种形式的核酮糖-1,5-二磷酸羧化酶/加氧酶激活酶对高温的敏感性不同。植物生理。114,439-444。
[16] Puhakainen T, Li C, Boije-Malm M, Kangasjärvi J, Heino P & Palva ET.(2004)。在银桦冷驯化过程中,低温诱导c -重复结合因子控制基因表达的短日增强。植物136, 4299 – 4307
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: CORNIC Gabriel (2025年1月5日), 温度对光合作用的影响, 环境百科全书,咨询于 2026年7月15日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/effects-temperature-on-photosynthesis-2/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。


