


Symbiosis and parasitism
PDF
Living organisms are permanently closely associated with each other. Their interactions can be classified according to the level of association of the organisms involved, the duration of these interactions and their beneficial (or not) impact on both partners. All intermediate situations exist, forming a true continuum from free organisms that need other organisms to feed themselves to parasites which life cycle is entirely based on specific hosts. Symbiosis and parasitism illustrate -beyond the extreme diversity of situations- that interactions are in all cases essential to partners’ lives, and are often at the origin of the emergence of new properties for the systems thus constituted. This is the case, for example, of microbiota associated with each of the living organisms. But it is also the case for organisms modified by parasites that infect them and even disturb the behaviour of infected hosts compared to healthy individuals.
1. Some definitions
The network of interactions and interdependencies that exists between billions of organisms within the biosphereA living space where all the Earth’s ecosystems are located, corresponding to the thin layer of the atmosphere, hydrosphere and lithosphere where life is present. This dynamic living space is maintained by an energy supply (mainly due to the sun) and the metabolism of living organisms in interaction with their environment.; a level of organization that is founder of the concept of biodiversity (read What is biodiversity?). These interactions are most often of mutual benefit and their role in the physiology and adaptation of organisms to the environment is essential. For example, many animals cannot digest without the help of bacteria in their digestive tract, most plants can only use the soil with fungi colonizing their roots, which they feed in return [1].
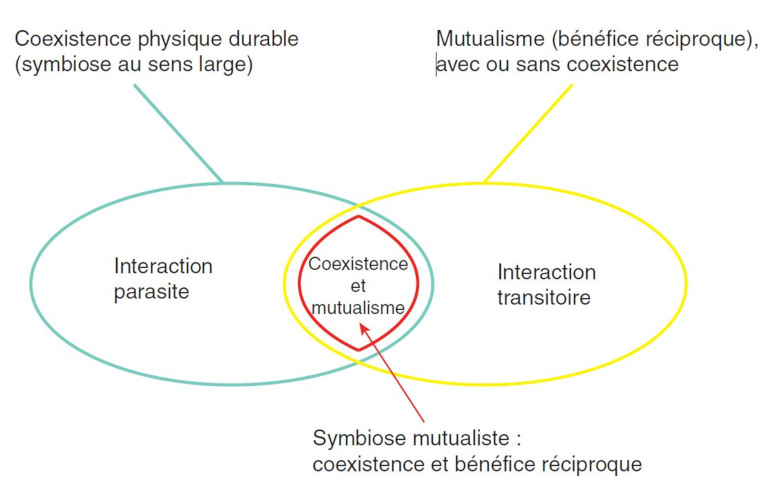
But this is not always the case: interactions between two organisms can be classified according to their beneficial, harmful or neutral effect for both partners. Thus, one can distinguish interactions that are beneficial for one partner and harmful for the other (predation, parasitism), beneficial for one and neutral for the other (commensalism) and mutually beneficial interactions (mutualism). In addition, all intermediate situations exist, in a true continuum of interaction types (Figure 1) [2]. They can also be classified according to their instantaneous (predation) or sustainable nature (parasitism, mutualism, etc.), as well as according to the degree of association between the partners [2]. Etymologically, the term symbiosis refers to “the common life of organisms of distinct species”. This broad definition refers to a sustainable coexistence, involving all or part of the life cycle of the two organisms, regardless of the exchanges between them. A more restrictive definition reserves the term symbiosis for sustainable and mutualist coexistence (see red part in Figure 1).
2. Mutualist symbiosis


Other benefits depend on the ability of one of the partners to move (pollination by bees, seed dispersal by ants or birds). On the balance sheet, similarly functioning associations have been set up several times during the evolution. Such convergences are illustrated by the diversity of insects cultivating fungi (ants, termites, beetles) and eukaryotesUnicellular or multicellular organisms whose cells have a nucleus and organelles (endoplasmic reticulum, Golgi apparatus, various plasters, mitochondria, etc.) delimited by membranes. Eukaryotes are, together with bacteria and archaea, one of the three groups of living organisms. that harbour photosynthetic algae in their cells (such as the appearance of chloroplastsOrganites of the cytoplasm of photosynthetic eukaryotic cells (plants, algae). As a site of photosynthesis, chloroplasts produce O2 oxygen and play an essential role in the carbon cycle: they use light energy to fix CO2 and synthesize organic matter. They are thus responsible for the autotrophy of plants. Chloroplasts are the result of the endosymbiosis of a photosynthetic prokaryote (cyanobacterium type) in a eukaryotic cell, about 1.5 billion years ago. in the eukaryotic cell) (see Symbiosis and evolution). All the organizations have had the opportunity to contract, during their evolution, one or more mutualist symbiosis(s). This is particularly true for large multicellular organisms, which constitute an ecosystem for microscopic organisms. The rhizosphere (the soil surrounding the root of plants) or the digestive tract of animals are thus major microbial niches, populated by thousands of species for each individual host, some of whose occupants are favourable to the host. As a result, each organism has a procession of symbiotes, especially developed in multicellular organisms.
3. Emerging symbiosis properties
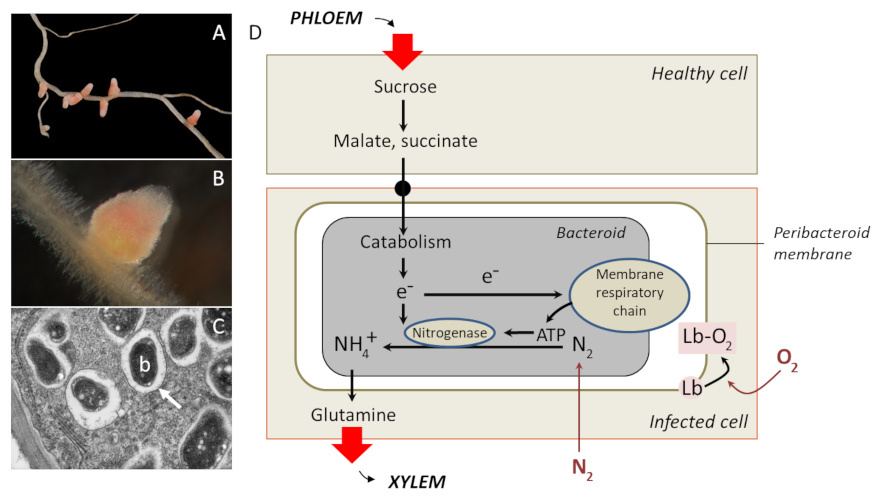
Other emergences are functional. In the example of nodules (Figure 4D), the bacteroid uses energy obtained from its respiration to reduce -thanks to the nitrogenaseEnzyme complex specific to certain prokaryotes that catalyzes the complete sequence of reactions during which the reduction of dinitrogen N2 leads to the formation of ammonia NH3. This reaction is accompanied by hydrogenation.– the atmospheric nitrogen N2 to ammonium NH3, which serves as a source of nitrogen for the plant (and bacteroid). Conversely, the plant provides carbon and oxygen supply. Oxygen is required for respiration, but nitrogenase is inactivated by oxygen: this contradiction explains why a free rhizobiumAerobic soil bacterium that can create symbiosis with legumes. These bacteria are found in nodules where they will fix and reduce atmospheric nitrogen, which can then be assimilated by the plant. In exchange plants provide carbonaceous substrates to bacteria. in the soil is unable to fix nitrogen. On the other hand, in the nodosity, oxygen does not diffuse freely, but is captured by a protein of the host cell, leghaemoglobin [7]. Located around the bacteroid, leghaemoglobin protects the nitrogenase from the inactivating effects of the oxygen and provides an oxygen reserve for bacteria respiration. Nitrogen fixation can therefore only be achieved within in the nodosity.
Many other functional traits are induced by symbiosis, such as some protective effects that rely on the induction of partner defences, tolerated by the symbiont but harmful to pathogens. Mycorrhizal fungi, for example, induce the accumulation of protective tannins at the root level, which are responsible for inducing an increased level of defence and reactivity throughout the plant, including the aerial parts. Thus, the mycorrhized plant reacts faster and more strongly to an herbivore or parasite than a non-mycorrhized control plant. In lichens, algae induce the fungus to synthesize secondary metabolites that have a protective role against strong light and herbivores.
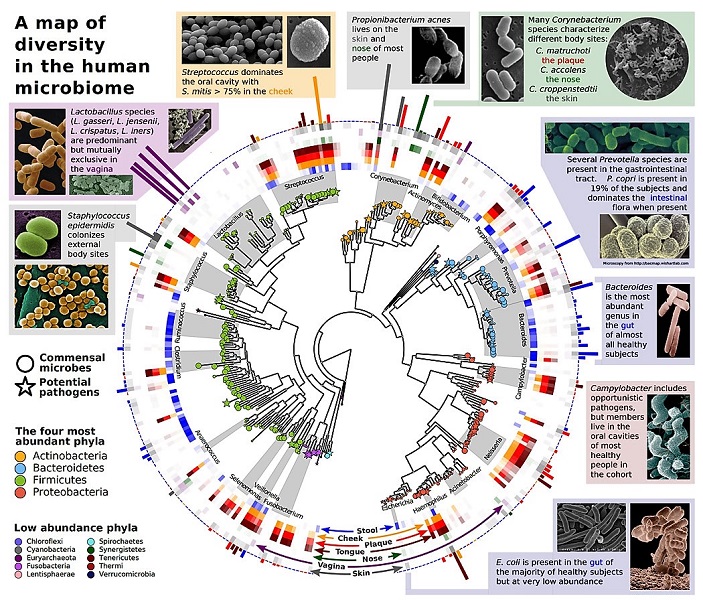
The gut microbiotaAll microorganisms (bacteria, yeasts, fungi, viruses) living in a specific environment (called microbiome) in a host (animal or plant). An important example is the set of microorganisms living in the intestine or intestinal microbiota, formerly called “intestinal flora”. is essential for the proper functioning of its human host, not only in terms of digestion or vitamin production, of course, but also for metabolism, immunity… or the nervous system. The imbalances in the intestinal flora are now suspected of being at the origin of a series of pathologies: obesity, diabetes, cardiovascular diseases, allergies, inflammatory diseases, even autism [2],[7]. The human microbiota is not limited to the digestive tract: international metagenomic programs have identified genes from a large number of symbiotic microorganisms living in the mouth, nose, vagina or on the skin (Figure 5).
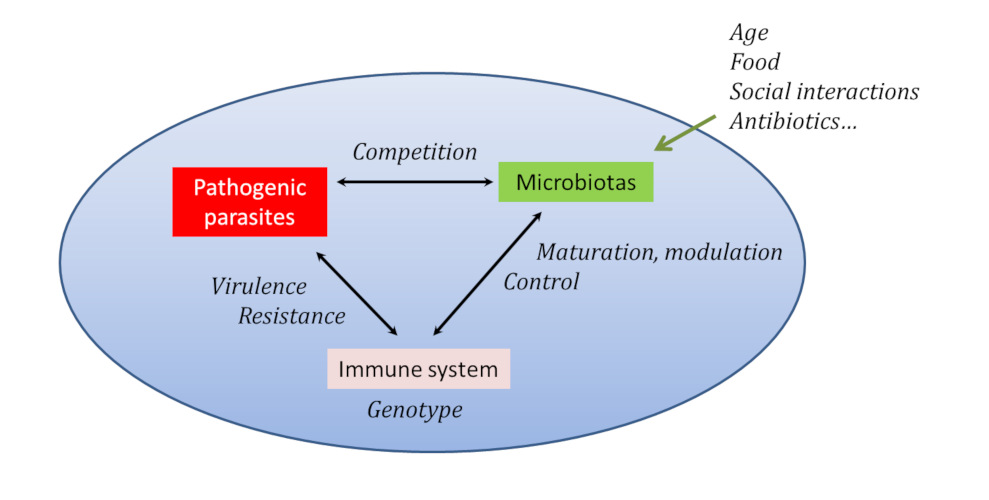
It has therefore been proposed that the unit relevant for biology or evolution should be less the organism than the symbiotic procession: we speak of holobiontemeans the biological unit composed of the host (plant or animal) and all its microorganisms. to name this entity more relevant to the importance of biotic interactions [11].
4. Parasitism, an evolutionary success story
If one of the partners in the symbiosis discovers how to use the other effectively, it becomes a parasite. There is indeed a continuum between symbiosis and parasitism [5]. The parasite exploits resources provided by another unrelated individual, the host, to the detriment of the latter. Parasitism is a long-lasting interaction with a host, unlike predation, where the interaction lasts only as long as the time of capture and digestion. However, from an evolutionary point of view, it can be said that predation is only an extreme form of parasitism. There are parasites that slowly kill their host. This is the case of plant parasitic fungi (Mildew, Armillaries, Hoof fungus, etc…) that complete their life cycle on dead tissues. When a cheetah grabs an antelope, there is an exchange of energy and only energy. In host parasite systems where the host survives (referred to as biotrophic parasitism), the duration of the interaction is quite different: the two organisms then live together, often one in the other, sometimes cell in cell or even genome within genome. The genetic information of each partner is expressed side by side and durably in a tiny portion of space [11].
All living beings are affected by parasitism as hosts or parasites (Figure 7). Among the known species, 30% of the approximately 2 million eukaryotic species are thought to be parasites [12]. The best known parasito-faunaAll the parasitic fauna of an organism. is that of man. It consists of 179 species of parasites, 35 of which appear to be specific to Homo sapiens [13]. This image can be increased by hyperparasitism (parasite’s parasites), a widespread phenomenon in parasitic arthropods and parasitoidsAn organism that develops on or in a “host” organism in a two-phase process: it is first biotrophic and then predatory, leading to the final death of the host. [14]. Recent estimates suggest that the world of viruses, which parasitize cells by diverting their functioning to the production of new viral particles, has been profoundly underestimated. They are present in all ecosystems and would constitute the most abundant and diversified genetic entities in living organisms [15].
For evolutionistsPartisans of evolutionism, who believe that species evolve over time,, host-parasite models raise countless questions about parasitism itself, the evolutionary dynamics of their interactions, and the evolutionary consequences on host species. The role played by parasites on the world of free species is indeed enormous. The success of the parasitic lifestyle has never been denied throughout the evolutionary process because a host offers, to any organism that knows how to exploit it, not only habitat and food but also an effective means of dispersal. While in the past, research has focused on the direct effects of pathogens on the fertility and survival of their hosts, current research illustrates consequences on such diverse traits as behaviour, selection processes and life history, to name but a few.

5. Parasitic cycles
The parasite cycle is the result of the transformations undergone by a parasite during its lifetime to ensure its reproduction, in the various ecological niches it occupies: host(s), external environment. While many parasite species have simple cycles, exploiting a single host species, others successively exploit several host species: this allows seasonal relays, or to multiply infectious forms, because the success rate of host colonization is often low. The complexity of the cycles has appeared several times independently during the evolution. Among the most complex records, we can mention the case of the trematode Halipegus ovocaudatus whose cycle includes 4 obligatory hosts: a mollusc, a crustacean copepod, a dragonfly larva and a frog. In addition to these extreme situations, complex cycles with two or three hosts are found, particularly in helminthsGeneric term that includes various types of worms that are generally parasitic: roundworms (nematodes), thorny trunk worms (acanthocephalus – “thorny-headed” worms) and flatworms (plathelminths : these are codes and trematodes). or rust (pathogenic fungi). In addition to the complexity of simple cycles, there are also complex cycles during the evolution of secondary simplifications.
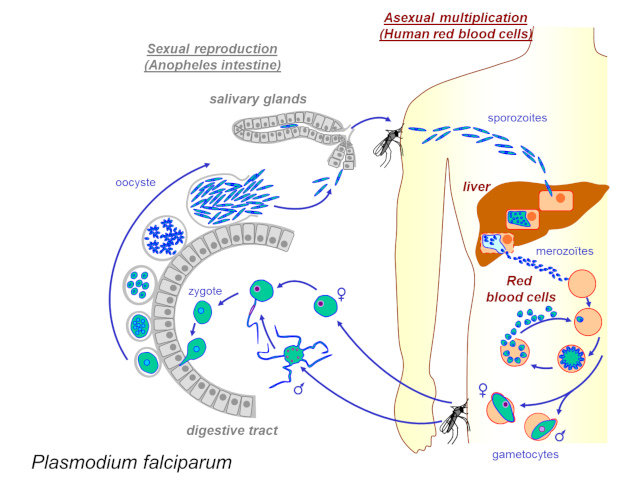
When infected with plasmodium, mosquitoes change their behaviour: they become more active, more aggressive and bite more people, thus increasing their probability of transmission [2]. These changes seem to be synchronized with the development of the parasite (e.g. decrease in mosquito bite rate when the parasite is immature, and increase when the parasite has reached the transmissible stage). Once in the vertebrate host, these same parasites seem to be able to modify the odours of the hosts to make them more attractive to mosquito vectors. This change in host behaviour after infection is a characteristic example of parasitic manipulation [2].
6. Parasitic manipulations
Some parasites are capable of significantly modifying the physiology, morphology or behaviour of their host with the consequence of increasing their probability of transmission. This host exploitation strategy is now described in many host parasite systems phylogeneticallyAdverb describing the result of an analysis of the relationship relationships between distant living things. distant. Phenotypic Characterize a trait or character of a living organism (anatomical, physiological, molecular or behavioural aspects), which can be analyzed. changes in infected hosts are generally considered an illustration of the extended phenotype concept [18]. These phenotypic changes actually correspond to the expression of the parasite’s genes and the effect of the corresponding proteins on the host’s phenotype. According to this idea, these induced modifications are adaptive for the parasite and not for the host.

Some parasite manipulations lead the host to suicidal behaviour. A well described case is that of non-segmented nematomorphsWorms with cylindrical bodies, extremely long and thin (on average from 0.5 to 2.5 mm in diameter for 10 to 70 cm in length). Also called Gordian worms because of the impression they give of making complicated knots with their bodies. worms, whose adult form lives in water and looks like a kind of thread. The host is usually a terrestrial insect, such as a grasshopper (Figure 7H) that hosts the larval form. The adult worm must return to the aquatic environment to reproduce. To do this, it manipulates the host’s behaviour, forcing it to jump into the water. Thanks to this final drowning, he can then return to the environment in which he completes his life cycle. However, this type of suicidal can be beneficial to noninfected animals of the same species, as it reduces the risk of contamination. This is the case of the ant parasitized by Ophiocordyceps (Figure 9B), which is then recognized as such and rejected from the anthill by its congeners.
In a few cases concerning plants, the determinism of handling is a little better known. It reveals a surprisingly convergent mechanism in pathogenic fungi and bacteria, but also in plant-parasitic nematodesRound, non-segmented worms. Some lead a “free” life (in soil, water, etc.). Others have a parasitic life, within fungal, plant or animal organisms.. They cause root deformations where they take shelter and feed, called galls. The genomes of these organisms encode a multitude of small secreted proteins (or peptides), which modify the functioning of other host proteins. We talk about effectors: some penetrate the host cells, and reorganize the metabolism or alter the defense reactions… Sometimes they act at the level of the cell nucleus and are responsible for changes in gene expression. It is likely that these mechanisms also play a role in other types of parasitism: they are even found in mycorrhizal fungi. This suggests that secreted peptides could contribute to the changes observed in mutualist symbiosis – again highlighting the existence of similarities in mechanisms between mutualist and parasitic symbiosis.
Beyond the spectacular and fascinating nature of parasitic manipulation, some of the pathogens involved are responsible for many crop losses, but also for serious diseases, including vector-borne diseases such as malaria mentioned above, dengueMosquito-borne viral infection in tropical and subtropical regions around the world. Causes a flu-like syndrome that can progress to life-threatening complications. There is no specific treatment for dengue fever,, trypanosomiasisInfections caused by trypanosome parasites, or leishmaniasisParasitic diseases causing very disabling or even fatal skin or visceral diseases if not treated. They are caused by various parasites of the genus Leishmania, transmitted by the bite of insects commonly known as sandflies., and thus represent major public health problems [19].
References and notes
Cover photo: © Jenny Huang from Taipeh (CC BY 2.0) via Wikimedia Commons.
[1] Selosse M.A. (2000). La Symbiose. Vuibert, Paris.
[2] Lefèvre T., Renaud F., Selosse M.-A., Thomas F. (2010). Chapitre 14, Évolution des interactions entre espèces, in F. Thomas, T. Lefèvre & M. Raymond (ed.), Biologie évolutive, p. 555-653. De Boeck, Paris.
[3] Selosse MA, Gilbert A (2011) Des champignons qui dopent les plantes. La Recherche 457, 72-75.
[4] Selosse MA & Gilbert A (2011) Mushrooms that boost plants. Research 457, 72-75.
[5] Cleveland A., Verde E.A. & Lee R.W. (2011) Nutritional exchange in a tropical tripartite symbiosis: direct evidence for the transfer of nutrients from anemonefish to host anemone and zooxanthellae, Marine Biology, 158: 589-602
[6] Corbara B http://www.especes.org/#/1-menage-a-trois/4541755
[7] Leghaemoglobin. Oxygen binding protein with a structure very similar to blood hemoglobin. Present in the nodules of legumes, it protects the enzymatic complex (nitrogenase/hydrogenase) from the effects of oxygen which inactivates it and constitutes a reserve of oxygen for bacteria (aerobic activity).
[8] Dawkins R. (1982) The extended phenotype. Oxford University Press, Oxford.
[9] Morgan X.C., Segata N. & Huttenhower C. (2013) Biodiversity and functional genomics in the human microbiome, Trends Genet. 29, 51–58
[10] Gross R., Vavre F., Heddi A. & Hurst G.D.D., Zchori-Fein E. &Bourtzis K. (2009) Immunity and symbiosis. Molecular Microbiology 3, 751-759.
[11] Selosse M.A. (2016) Au delà de l’organisme : l’holobionte. Pour la Science, 269, 80-84.
[11] Combes C. (1995) Interactions durables. Écologie et évolution du parasitisme. Éditions Masson, 525 p.
[12] From Meeûs T. & Renaud F. (2002) Parasites within the new phylogeny of eukaryotes. Trends in Parasitology 18, 247-251.
[13] De Meeûs T., Prugnolle F. & Agnew P. (2009) Asexual reproduction in infectious diseases. In Lost Sex, Schön I, Martens K & van Dijk P eds, Springer, NY, p. 517-533.
[14] Parasitoid: an organism that develops on or in a “host” organism in a two-phase process: it is first biotrophic and then predatory, leading to the final death of the host.
[15] Hamilton G. (2008) Welcome to the virosphere. New Scientist 199, 38-41.
[16] Source http://www.archive.org/stream/traitzoologiqu00brem#page/n613/mode/2up
[17] Zheng et al (2011) Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine. Genome Biology 12: R116
[18] Dawkins R. (1976) The selfish gene. Oxford University Press.
[19] Lefèvre T. & Thomas F. (2008) Behind the scene, something else is pulling the strings: Emphasizing parasitic manipulation in vector-borne diseases. Infection, Genetics and Evolution 8, 504-519.
The Encyclopedia of the Environment by the Association des Encyclopédies de l'Environnement et de l'Énergie (www.a3e.fr), contractually linked to the University of Grenoble Alpes and Grenoble INP, and sponsored by the French Academy of Sciences.
To cite this article: SELOSSE Marc-André, JOYARD Jacques (January 5, 2025), Symbiosis and parasitism, Encyclopedia of the Environment, Accessed July 5, 2026 [online ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/en/life/symbiosis-and-parasitism-2/.
The articles in the Encyclopedia of the Environment are made available under the terms of the Creative Commons BY-NC-SA license, which authorizes reproduction subject to: citing the source, not making commercial use of them, sharing identical initial conditions, reproducing at each reuse or distribution the mention of this Creative Commons BY-NC-SA license.


