




什么是生物多样性?

生物多样性涉及所有的生物体、生物体之间以及生物体与环境之间的相互作用,涉及生命的各个层次。从基因到个体,再到物种和种群,到生态系统中的群落。生命之树说明了物种的多样性,反映了物种之间的关系,帮助我们认识各个物种的进化历史。因此,从广义上讲,生态系统的特征是各组分之间的相互作用、物质和能量流动,以及随着时间的推移,可持续性与演化、恢复力、抵抗力间之的动态平衡。
“重要的东西是眼睛看不见的。”小王子又说了一遍,这样他就能记住了。
——《小王子》21章 安托万·德·圣埃克苏佩里
1. 定义
生物多样性的概念是最近才提出的。1984年,爱德华·O·威尔逊在其《生物多样性》一书中首次提出了生物多样性的概念,但这个新概念真正开始流行是在1992年里约地球峰会《生物多样性公约》签署之后。该公约的第2条对生物多样性做了这样的定义:“所有来源的形形色色生物体,这些来源包括但不限于陆地、海洋和其他水生生态系统,以及它们所构成的生态综合体;这包括物种内部、物种之间和生态系统(生态系统由生物(或生物群落)及其生物、地质、土壤、水文、气候和其他环境(生物群落)组成。生态系统的特点是生物物种与其周围环境之间的相互作用,生态系统各组成部分之间的物质和能量流动,使其得以生存,并随着时间的推移在可持续性和演化之间保持动态平衡。)的多样性。”生态学家罗伯特·巴尔博将这个定义总结为“万象生命”。
因此,生物多样性涉及所有生物体、生物之间以及生物体与所处环境之间的相互作用;生物多样性的范围涵盖了生命系统的全部层次:从基因到个体,再到物种,它们都与所处的环境、周围的物种、尤其与生活其间的生态系统发生着密切的相互作用。我们还必须从地球历史的角度来理解生物多样性:大约38亿年前地球上出现了生命(见《生命的诞生》系列文章),当前生物的多样性正是漫长时间演化的结果。
然而,生物体的生物和遗传资源及其生存环境的可持续性是社会、经济、法律问题,与人类社会和整个生物圈息息相关,包括资源的获取、利用、获益、共享、管理、可持续性,等等。最后,生物多样性还是一个伦理问题,比如环境伦理学(见《聚焦环境伦理学》)等几种活跃的哲学流派所捍卫的不受时效限制的物种生命权。因此,正如雅各·布朗德尔(Jacques Blondel)在《生物多样性不是奢侈品,而是必需品》中所描述的,生物多样性也在人文社科(Human and Social Sciences)中有其一席之地。
2. 物种多样性
在生物多样性的层次中,物种多样性看上去最为直观、易于理解,因为物种多样性区分了物种。我们很容易就能辨认周围的各种动植物:我们知道什么是百合、蜘蛛、企鹅或豹子(图1)。但是物种的定义就没有这么简单了。动物学家和系统学家(研究分类学的生物学家,使用一种系统来计算生物,尤其是根据逻辑原则将生物按照一定的顺序排列,从而对它们进行分类。)继尧姆·勒克莱特(Guillaume Lecointre)认为:“自然界中并没有物种,只有生殖隔离。我们根据理论模型创造了物种。”[1]简而言之,物种就是一群外形相似,在自然条件下可以交配,并能生育出能够存活的可育后代的生物。然而,这个定义并不适合如细菌这样的微生物,它们肉眼不可见,很难用简单的形态学标准来区分。

这些物种是根据林奈提出的双名命名法命名的;后面是发现者的名字(L.代表林奈),所标注的年份是这些生物被描述的年份。[来源:照片由© Jacques Joyard惠赐]
地球上有多少种生物?目前估计约有三百万到一亿个物种,其中已被发现、描述的物种只有大约一百七十万到两百万个。显然,描述得最好的是我们能够直接接触到的物种:陆地植物——总数估计有三十万,其中二十多万已被描述——和脊椎动物,尤其是鸟类。鸟类总数约有十万,虽然人类已经描述了其中几乎99%的鸟类,但是每年都会有新的鸟类物种被发现!另一方面,微生物中只有1%被描述,如病毒,古细菌(生活在极端环境(厌氧、高盐度、酷热……)中的单细胞原核微生物。卡尔-沃斯(Carl Woese)和乔治-福克斯(George E. Fox,1977年)的系统发育研究将古细菌与其他原核生物(细菌)区分开来。目前,生物被认为包括三类:古菌、细菌和真核生物。)、细菌等等。因此,这些生物是许多密集的研究项目的重点,比如2009年到2012年间,塔拉海洋计划(Tara Oceans)考察队环绕地球航行,对海洋中的浮游生物(水生环境(海洋、湖泊……)中随水流漂浮的所有生物微生物。通常肉眼看不见,大小从0.2微米(0.002毫米)到0.2毫米不等。浮游生物分为植物浮游生物和动物幼苗浮游生物。)物种进行了统计[2]。研究人员收集了全球主要的海域的病毒、微生物和真核生物(单细胞或多细胞生物,其细胞具有细胞核和细胞器(内质网、高尔基体、各种质体、线粒体等),并由薄膜分隔。真核生物与细菌和古细菌是三类微小生物之一。)(从单细胞藻类到鱼类幼苗),获得了超过35,000种不同的浮游细菌的遗传物质,这些细菌大部分至今仍不为人所知。
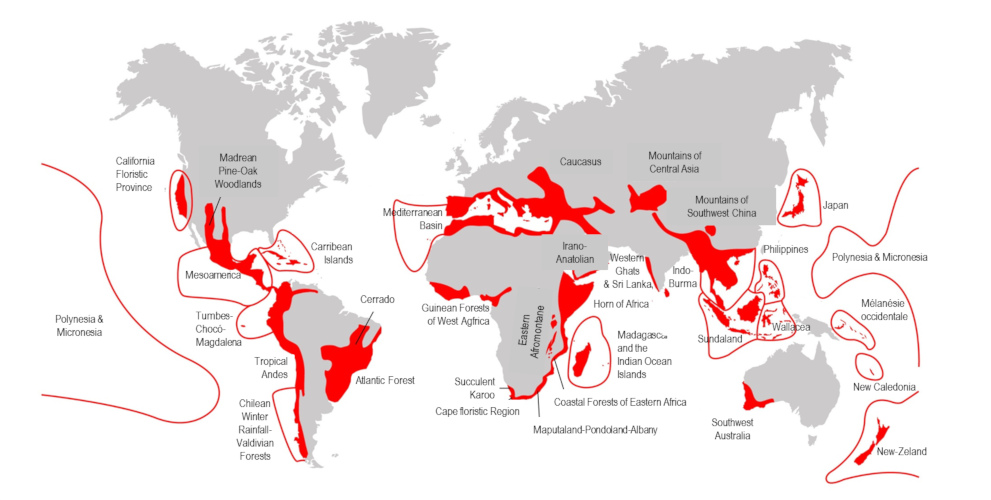
[来源:改编自©保护国际(2005年2月)]。
描述现存或灭绝的物种对物种名录编制和物种分类至关重要。最初,根据不同的地区和当地语言,每个物种都有各自的俗名。瑞典植物学家、医生和动物学家卡尔·冯·林奈[3]提出了双名命名法(动植物物种的科学命名模式,包括在属名后面加上物种名称的限定词。),使特定物种得以精确命名。18世纪提出双名命名法时,人们认为物种是由形态学标准定义的固定实体。卡尔·冯·林奈根据花的结构对植物进行分类,更确切地说,是根据生殖器官(雄蕊和雌蕊)的数量、排列和比例进行分类。到了19世纪,物种不变不灭的理念被终结。最开始是乔治·库维耶[4](见《焦点:乔治》)意识到一些动物曾经存在过,但如今已不复存在:之前无法解释的大量化石就是包括从贝壳到恐龙在内已经灭绝的物种(见《焦点:古生物学家的物种》)。
所有这些发现促成了生物体的经典分类体系(或分类等级)的形成,这一体系以可观测的特征为基础,构建层级分类结构,即(生物)→域→界→门→纲→目→科→属→种。
达尔文提出自然选择导致物种演化(见 进化论和焦点:达尔文),也就是说物种间具有亲缘关系,由共同的祖先演化而来,自然选择学说彻底挑战了这种层级结构。 农民或牧民会在每一代作物或牲畜中选择具有最优良特性的个体进行培育,达尔文类比这种人工选育,提出了“自然选择”这一术语。这一革命性的概念会使我们有可能思考一个公认的事实:在同一物种中,相对而言一些个体互为相似,但所有的个体都各不相同。我看起来像我的父母、兄弟或姐妹,但我与他们不同。
自20世纪下半叶以来,系统发育分类法(见下文)就沿着演化的观点发展。人们用系统发育分类法来说明解释物种之间的亲缘关系远近,从而了解它们的演化历史或系统发育(个体或生物群(如物种或种群)之间的进化关系的研究。)。
3. 种内多样性、遗传多样性和物种演化

[来源:上层:© Jean-Claude Melet(见参考文献[5]);下层:© Catherine Lenne(见参考文献[6])
达尔文认为,物种的每一代都是由具有相似特征、但在环境中的生存能力又各不同的个体组成的,每一个个体都是一系列性状(物理、遗传、适应环境的能力……)的综合体。在面对环境(气候、捕食者、寄生物、资源等)的变化和制约,有些个体难以生存或繁殖,最终会从群体中消失;而其他能更好适应和存活的个体会将优势的特征遗传给后代。
生活在一个相对较小的地理区域内的同一物种的所有个体,具有基因上的亲缘关系,但又各不相同,这就是一个种群。当出现地理隔离时,如果环境条件适宜,种群内每个个体就会或快或慢地发生改变,经过一代又一代后,就会形成与原来种群不一样的特征或技能,这就是同一物种内的多样性或者说种内多样性。这种分化最终可能导致新的个体不能与原有种群中的个体进行有效繁殖,于是一个新的物种就诞生了(图4)。达尔文雀(见焦点:)就是一个典型的例子。华莱士[7](Alfred R. Wallace)在19世纪就已经想到了地理环境在物种形成(从原始物种的种群中分化出来的新物种进化过程。)中的重要作用,而1915年阿尔弗雷德·魏格纳[8]提出大陆漂移假说(是当今板块构造学[9]的原型)时,就利用到了这一点。魏格纳发现二叠纪(两亿年前)之前美洲和非洲大陆上的一些动植物化石非常相似,二叠纪之后每个大陆上的化石开始变得不同。魏格纳于是想象各大陆是由一个超级大陆——盘古大陆(是在石炭纪时期由地球表面现有大陆的碰撞形成的,然后重新组合了所有出现的陆地,在三叠纪分裂为两个大陆:北部的劳亚大陆和南部的冈瓦纳大陆。)——分裂产生的。一个典型的例子是平胸鸟类:如今鸵鸟生活在非洲和南美洲,鸸鹋在澳大利亚,几维鸟在新西兰,但这些鸟类的祖先都是遍布盘古大陆的”古鸵鸟”。
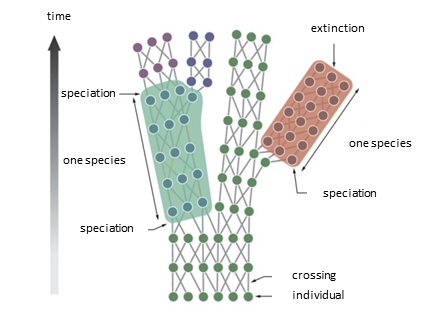
[来源:根据Lecointre(2009)[见参考文献10]。(time:时间;speciation:物种形成/成种;one species:一个物种;extinction:物种灭绝;crossing:遗传信息交换;individual:个体。)
在达尔文提出其理论的同时,孟德尔[11]发现了生物有机体的特征是如何代代相传的。这一发现在当时并未引起关注,但却开启了遗传学的大门,使我们得以了解物种演化的起源机制。到20世纪上半叶,遗传学才逐渐与达尔文理论结合。然而,直到1953年,沃森和克里克发现了DNA(脱氧核糖核酸)[12]的结构,并揭示了其功能之后,人们才认识到DNA起着遗传的载体和生物体后代标记的作用,由此引发了对生物体基因组性质(见介于稳定性和可变性之间的基因组),以及导致突变和重组机制的全面研究。突变和重组不仅对生物个体产生影响,也对种群适应环境变化(即演化)至关重要。种内多样性实际上就是遗传多样性。基因突变引起的变异是个体多态性(多态性与基因突变引起的变异相关,是指群体中基因 DNA 核苷酸序列的变异,也指动物、植物、真菌或细菌群体中给定基因或基因座的多个等位基因的共存)的来源(见遗传多态性与选择)。从自然选择的角度来看,这些突变可能是 “中性的”、“轻微有害的” 或 “有利的”。由于适应性的不同,这些突变在物种或种群的基因库或得以保留,或被淘汰。因此,每个物种都有独特的基因组合,同时物种的每个个体都会有区别于该物种其他个体的特征。
即使是同卵双胞胎也会表现出性状的差异,这是由于生物体内还存在一个更加精细的调节,即表观遗传调节(见表观遗传学,基因组及其环境)。表观遗传调节是指不通过DNA编码序列变化,而通过对DNA进行生物化学修饰(例如甲基化)的方式进行的调节。因而,同一段遗传编码在生物发育过程中可以表达出不同的产物。有些表观遗传标记很短暂,但也有一些标记能长期存在,甚至可能传递给后代,不过一般不会长久地遗传下去。然而,由于表观遗传修饰没有改变DNA序列,没有改变种系的遗传结构,因此不会导致新物种形成(见适应:对环境挑战的反应)。
4. 生命之树
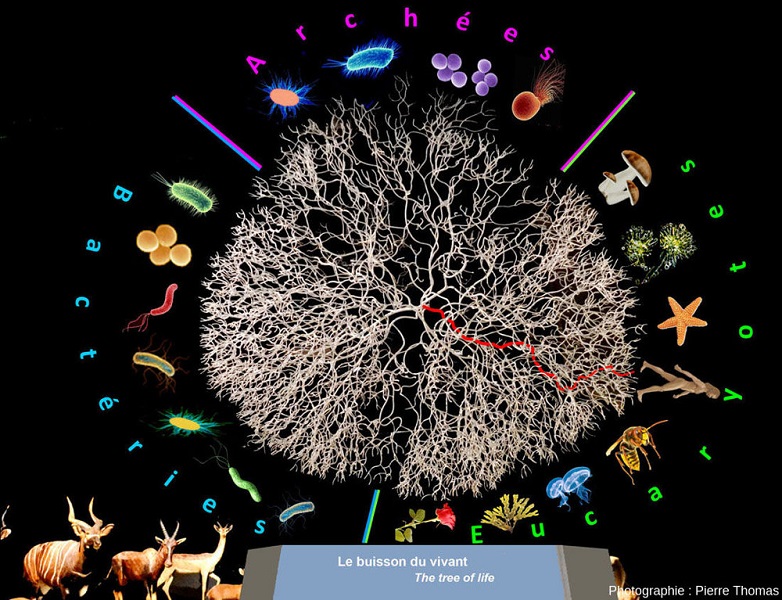
[来源:法国里昂汇流博物馆雕塑。© 2014 Samba Soussoko设计,Laetoli 制作;图片编辑:© Pierre Thomas, ENS Lyon](Bacteries:细菌;Archees:古细菌;Eucaryotes:真核生物)
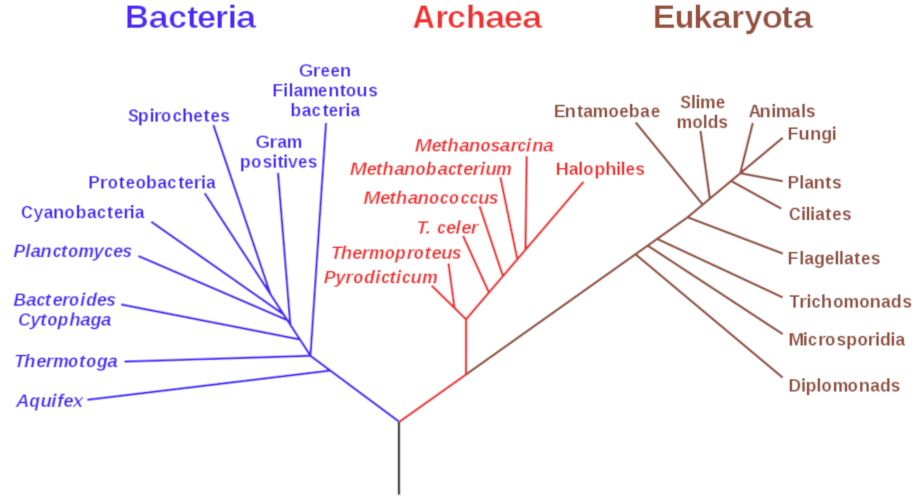
[来源:Eric Gaba,美国宇航局天体生物学研究所]。(Bacteria:细菌;Archaea:古菌;Eukaryota:真核生物;Aquifex:产液菌属;Thermotoga:热胞菌;Bacteroides:拟杆菌;Cytophaga:嗜细胞菌属;Planctomyces:浮霉状菌属;Cyanobaceria:蓝藻;Proteobacteria:变形菌;Spirochetes:螺旋菌;Gram positives:革兰氏阳性菌;Green Filamentous bacteria:绿色丝状细菌;Thermoproteus:热变形菌属;Methanococcus:产甲烷球菌属;Methanobacterium:甲烷杆菌属;Methanosarcina:甲烷八叠球菌属;Halophiles:噬盐菌;Entamoebae:变形虫;Slime Molds:黏菌;Animals:动物;Fungi:真菌;Plants:植物;Ciliates:纤毛虫;Flagellates:鞭毛虫;Trichomonas:滴虫;Microsporidia:小孢子虫;Diplomonads:双滴虫)
5. 生态系统
生态系统的经典定义是一个地点所有生物(或称生物群落)与群落生境(即生物、地质、土壤、水文、气候等)构成的整体。因此,生态系统的数量无穷无尽:泥炭沼泽(见泥炭地和沼泽,非凡的湿地)、森林、海底的“黑烟囱”(见黑烟囱的生态系统)是众所周知的生态系统;而反刍动物的瘤胃、卡门培尔奶酪或正在腐烂分解的生物体也构成不同的生态系统(见“生态系统“标题下的系列文章)。广义上讲,生态系统的典型特征是(生物物种之间及其与周围环境之间的)相互作用,以及各组分之间的物质和能量流动,从而使生命和生态系统能够随着时间的推移,在持续性与演化之间保持动态平衡。一个生态系统中的“物质”指的是在环境中所有的生物(植物、动物、生物)和非生物要素(空气、营养物质、水)。
5.1. 生态系统中物种的相互作用
同一生态系统中的生物之间存在相互作用、相互依赖的网络,这是生物多样性概念的核心本质(见共生和寄生)。这些相互作用通常是互利的,对生物的生理和环境适应至关重要。例如,许多动物如果消化道内没有细菌和古菌就无法消化食物;而大多数植物只能通过定殖于根部的真菌从土壤中获取资源,真菌反过来也从植物那里获得营养,这种真菌/根系的结合称为菌根共生。
但情况并非总是如此:两个生物之间的相互作用可以根据对双方有益、有害或中性进行分类。我们可以区分对一方有利而对另一方有害的相互作用(捕食,寄生)、对一方有利而对另一方中性的相互作用(偏利共生)或者对双方都有利的相互作用(互利)。即便现实中上述各种相互作用的中间类型都可能存在,形成真正的相互作用类型的连续的变化谱。我们还可以根据相互作用的瞬时性(捕食)或持续性(寄生、互利等)进行划分,或者根据生物之间相互作用的强度分类。从词源上看,共生一词指的是 “共同生活”,这明确指出共生是两种生物在整个或部分生命周期中存在持续的、专性的共存关系,无论它们之间的相互作用如何。狭义的共生则是指双方持续、互利的共存关系(见共生和寄生)。
因此,在一个生态系统中,成千上万的物种共存,形成了极其复杂的相互作用,这是生态系统功能的基础。生态系统中相互作用的变化会导致生态系统功能多样性发生动态演变,如提供生态系统服务(我们从生态系统中获得的好处无需采取行动即可获得。各种服务是生态系统功能和维护自然过程的结果。 因此,供应服务本身提供食物、水、木材和纤维等物品。调节服务负责调节气候和降水、水(例如洪水)、废物和疾病传播。 文化服务涉及美感、灵感和娱乐,有助于我们的身心福祉。辅助服务包括土壤形成、光合作用和肥料物质的回收,没有这些就没有生长或生产。)的能力变化(见生物多样性不是奢侈品,而是必需品)。
5.2. 生态系统中物质和能量的流动
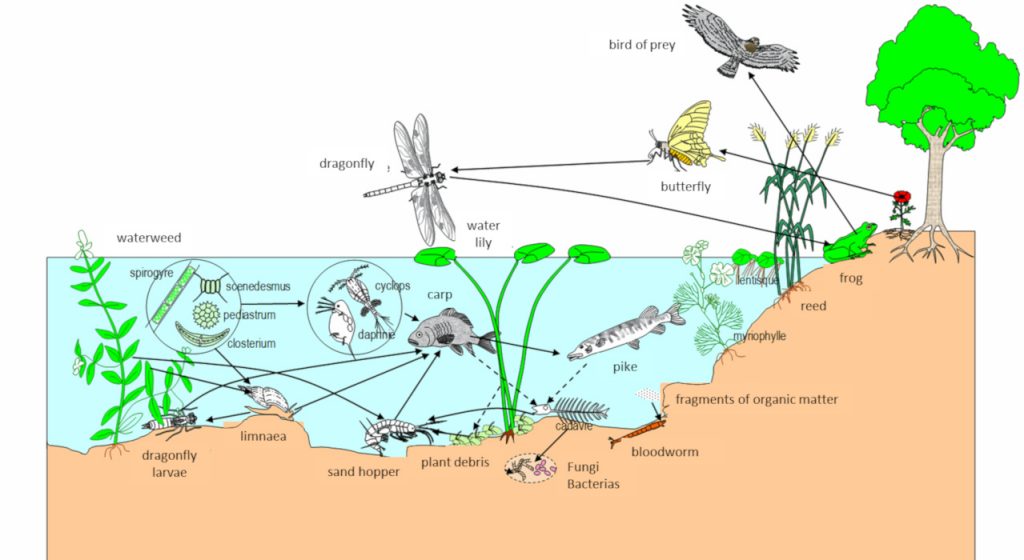
[来源:© Alain Gallien,第戎学院图示库(http://svt.ac-dijon.fr/schemassvt/)](bird of pray:猛禽;dragonfly:蜻蜓;butterfly:蝴蝶;water lily:莲;water weed:水草;spirogyre:水棉;scenedesmus:栅藻;pediastrum:盘星藻;closterium:新月藻;cyclops:剑水蚤;daphnia:水蚤;carp:鲤鱼;pike:狗鱼;reed:芦苇;myriophylle:狐尾藻;fragments of organic matter:有机物残屑;blood worm:红蚯蚓;cadaver:残骸;fungi:真菌;bacterias:细菌;plant debris:植物残片;sand hopper:沙蚤;dragonfly larvae:蜻蜓幼虫;limnaea:椎实螺)
物质遵循拉瓦锡(Lavoisier)所阐述的物质守恒定律:“物质不会凭空消失,也不会凭空产生,只会从一种形式转变为另一种形式”。在图7所示的例子中,一种植食性水生腹足类椎实螺属动物以各种水生植物和藻类为食,这些植物和藻类都是生活在这个池塘中的初级生产者(能够通过光合作用等方式从矿物质中产生有机物的生物。它们是自养生物,位于食物链的底层。它们被初级消费者摄入,本身也可能成为次级消费者的目标。)。椎实螺取食后只保留了一部分食物中的物质,它们几乎完全以自养生物为食,因此属于初级消费者(一种需要消耗其他生物来制造自身有机物以不断生长的生物,是一种异养生物。只食用陆生或水生植物的食草动物属于初级消费者。)。鲤鱼不仅取食碎屑,也捕食椎实螺等小动物,它们属于次级消费者(以本身是消费者的生物为食的异养生物。肉食性掠食者(狼、狮子等)通常就是这类消费者,它们只以其他动物为食,处于食物链的顶端。)。另一种可怕的食肉动物狗鱼(Esox)也是如此,它的猎物种类繁杂,包括各种鱼类、两栖动物、蜥蜴、幼鸭、啮齿动物……,将猎物的部分物质保留下来。在池塘周围,特别是岸边,食肉鸟类——如白腹鹞——能够轻而易举地搜寻到猎物:小型哺乳动物、青蛙、鱼、昆虫和其他鸟类。
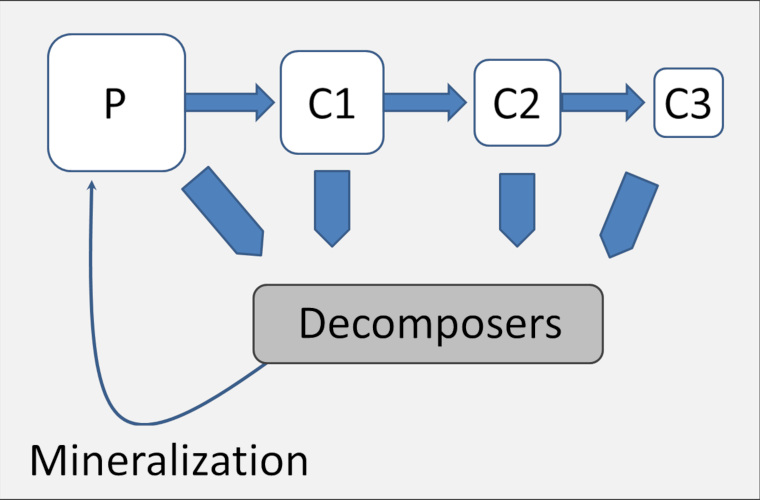
【来源:改编自 Online Academy [20]】(Decomposer:分解者;Mineralization:矿化作用)
同样,能量也在生态系统中流动。草地、森林或池塘生态系统的主要能量来源于太阳光能,通过自养生物[21]的光合作用将光能转变为化学能。植物吸收太阳光能,利用水、矿物质和大气中的二氧化碳制造有机物。通过食用这些有机物,消费者(异养生物,如如椎实螺、青蛙、狗鱼和白腹鹞)就可以回收植物产生的能量。这些能量有很大部分会流失:部分随着废物排出而损失,部分通过热量的形式散失,但其余的能量会被生物利用,使其得以生长、发育和繁殖。因此,能量通过食物网不停地流动。
能量总是从一个营养级流失到另一个营养级。由于能量在生态系统中不能循环利用,因而生态系统的运作需要从外界持续输入能量,如提供光合作用所需的光能。在另外一些生态系统中(如海底热液滋养的生态系统),自养过程需要的能量不是来源于太阳光能,而是由化学分子提供的,因而被称为化能合成作用(见极端环境中的微生物)。
5.3. 生态系统的动态
生态系统是在内、外因素作用下动态实体,其中气候和土壤类型等外部因素控制着生态系统的总体结构和功能。每个生态系统都有特定的环境条件(温度、湿度、pH值、土壤矿物组成等),使得植物、动物及微生物种群得以通过物质循环和能量流动而生存、相互作用和演变。反过来,生物物种随着时间的演化也改变了生态系统。生物与环境的复杂关系为各物种种群的生存提供了必要的条件和资源。土壤在陆地生态系统中是至关重要的,土壤不仅为初级生产者提供了固着的基础,为许多动物提供了多种多样的栖息场所(如洞穴),而且还是水和其他物质,特别是矿物质积累、转化和迁移的介质。
资源输入生态系统的过程通常受外部过程控制,但是资源的可及性是由生态系统内部因素决定,包括有机物分解、物种分布、根系间的竞争等。生态系统的生物环境(与生命有关。生态系统的生物因素是指植物群和动物群以及它们之间的关系。生命得以发展的环境。)和非生物环境(生态系统中影响特定生物群落的物理和化学因素。与生物因素相对,构成了该生态系统生态因素的一部分。气候因素(温度、光照、空气……)和化学因素(空气气体、矿物元素……)属于非生物因素。)存在着年际变化。严重干旱、特别寒冷(或温和)的冬天或害虫爆发,都会引发动物种群数量的巨大变化。在食物丰富的时候种群数量增加,而当食物稀缺时种群数量会锐减。因此,污染、干旱、温度变化以及疾病都会影响生态系统,栖息在生态系统中各种群的多样性的高低将决定生物能否在干扰的冲击下生存下来。
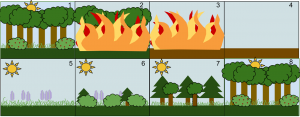
森林中的植物演替是个突出的例子。森林因疏伐、风暴或林火的作用而出现缺口,即林窗(图9)。演替循环从先锋(pioneering)阶段开始,向平衡阶段——演替顶级——发展,直到被另一个干扰(如火、暴风雨、洪水、滑坡、雪崩……)阻断,使演替循环又回到先锋阶段。在图9所示的例子中,森林被大火夷为平地,保留在土壤中的种子和被风、水和动物带来种子伺机而发。首先长出来的是称为先锋物种的禾草和其他草本植物,随后小灌木和树木幼苗将该地区重新覆盖。在这一阶段,针叶树快速生长,阔叶树长势较慢,被针叶树所荫蔽。随着高大的常绿或落叶树木冠层的发展,林分的郁闭度越来越高,不耐阴的物种随逐渐消亡,直到生态系统恢复到与此前类似的状态,这轮演替即告结束,待下一次干扰发生后又开始新一轮的演替循环。因此,随着一系列灾难性事件的发生,植物群落以及与之相关的微生物、真菌和动物群落群落也发生了渐次替代的演替过程。
事实上,经过百万年的演化和适应,生态系统发生了巨大的变化:石炭纪的泥炭沼泽可能与侏罗纪的山地泥炭沼泽毫不相干,但是控制这些沼泽的生态功能的机制很可能是一样的(见第一个复杂的生态系统和泥炭地和沼泽,非凡的湿地)。
如今,我们必须考虑一个额外的因素,即智人的影响(见生物多样性不是一种奢侈品,而是一种必需品)。事实上,我们对地球上的生物多样性构成了全方位的威胁。首先,因为农业活动的增强、过度捕捞和各种污染,的生态系统被分割、破坏或改变。土地转变为建造各类构筑物、道路、停车场、建筑物等的人类用地,也使生物多样性受到严重威胁。最后,运输和贸易的发展也破坏了生物多样性,尤其加重了如亚洲胡蜂等外来生物的入侵(见气候变化和全球化,昆虫入侵的驱动因素)。在上述因素的作用下,生态系统可能会完全丧失恢复力。更普遍的情况是,人类活动改变甚至严重破坏了自然环境,包括从景观格局(见阿尔卑斯山景观与生物多样性)到生物地球化学循环过程(见人类活动破坏的碳循环和环境中的硝酸盐)的方方面面。
致谢:凯瑟琳·伦内(Catherine Lenne)是位于Clermont-Ferrand的布莱斯帕斯卡大学(INRA et Université Blaise Pascal UMR PIAF)的教授,植物学知识渊博(对其它领域也非常熟悉),感谢她在本文撰写时提出的宝贵意见和建议。
参考资料及说明
封面照片:非洲水雉(Actophilornis africanus)行走在肯尼亚巴林戈湖中的睡莲叶上。[来源:Jacques Joyard]
[1] Lecointre G (2011) Les espèces, c’est nous qui les faisons! Espèces 1,68-72 (in french)
[3] Carl von Linné was know as Carl Linnaeus,1707-1778, before his ennoblement; Biography on Wikipedia
[4] Georges Cuvier,1769-1832; see focus Georges Cuvier
[5] http://www.florenum.fr/ (in french)
[6] http://www.desfleursanotreporte.com/ (in french)
[7] Alfred R. Wallace,1823-1913; http://people.wku.edu/charles.smith/wallace/BIOG.htm
[8] Alfred Wegener (1930); Wikipedia
[9] http://planet-terre.ens-lyon.fr/article/derive-continents-wegener.xml (in french)
[10] Lecointre G.(2009)Guide critique de l’évolution », Belin (in french)
[11] Johann Gregor Mendel (1822-1884); Wikipedia
[12] James Watson (born 1926) & Francis Crick (1916-2004); Biographies (as well as those of Rosalind Franklin,1920-1958,&Maurice Wilkins-2004) on the website: http://www.medecine.unige.ch/enseignement/dnaftb/19/concept/index.html
[13] Guyader H & Lecointre G (2013) La classification phylogenetique du vivant. Belin Paris,608p., ISBN 2701134560; http://www7.inra.fr/dpenv/leguyc46.htm
[14] Note that this ancestor remains hypothetical; he can only be identified by a few characters that he possesses and transmits to his descendants (the shared characters observed on the descendants), as puzzle pieces, but we do not have the complete “picture”.
[15] http://planet-vie.ens.fr/content/classification-vivant-mode-emploi (in french)
[16] It should be noted that the characteristics taken into account are so numerous and so heterogeneous that it is always necessary to choose, and depending on which ones are retained or eliminated, we end up with different cladograms, or trees. In this case, preference is given to the tree with the fewest knots (i.e. the most parsimonious).
[17] Woese CR & Fox GE (1977) Phylogenetic structure of the prokaryoticdomain: The primarykingdoms, Proc. Natl. Acad. Sci. USA 74,5088-5090.
[18] Hug LA et al (2016) A new view of the tree of life, Nature Microbiology 1,#16048; doi:10.1038/nmicrobiol.2016.48
[19] http://www.fondation-lamap.org/fr/page/10998/la-classification-des-tres-vivants-principes-g-n-raux (in french)
[20] http://www.academie-en-ligne.fr/Ressources/7/SN12/AL7SN12TEPA0013-Sequence-07.pdf (in french)
[21] Autotrophy: The ability of an organism to produce organic matter from the reduction of inorganic matter and an external energy source: light (photoautotrophy, as in the case of photosynthesis) or chemical compounds (chemoautotrophy).
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: JOYARD Jacques (2024年3月7日), 什么是生物多样性?, 环境百科全书,咨询于 2026年8月3日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/what-is-biodiversity/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。
