




植物固定的生命及其限制因素

植物扎根于土壤,土壤为它们提供了水分和矿物质,植物叶片通过吸收太阳能来固定二氧化碳中的碳。地球生命的基本过程依赖于这些静止的有机体从而运转起来。植物不可能像动物那样,通过不断的移动找到适合自己的栖息地,因此植物必须要适应环境中存在的各种差异和波动。从针叶林带到沙漠带,进化的力量推动了植物的发育和生理机能的完善,从而适应地球上热带或温带等不同的气候带。由此塑造出丰富的植物多样性。但是,地球上同一地点的环境也会随着季节的变化而变化。于是,同一物种的植物便形成细胞和分子机制,使得它们感知外部变化,然后重新排列基因表达。因此,它们可以重新调整自己的发育、生理和新陈代谢,以便尽可能有效地适应这些变化。
1.植物:适应而非移动的固着生物
“适应”的英文单词“adaptation”这个词来源于拉丁语adaptare。它可以被定义为生物在行为、生理或结构上的一系列调整或改变,从而使其能够更好地在特定的环境中生存。

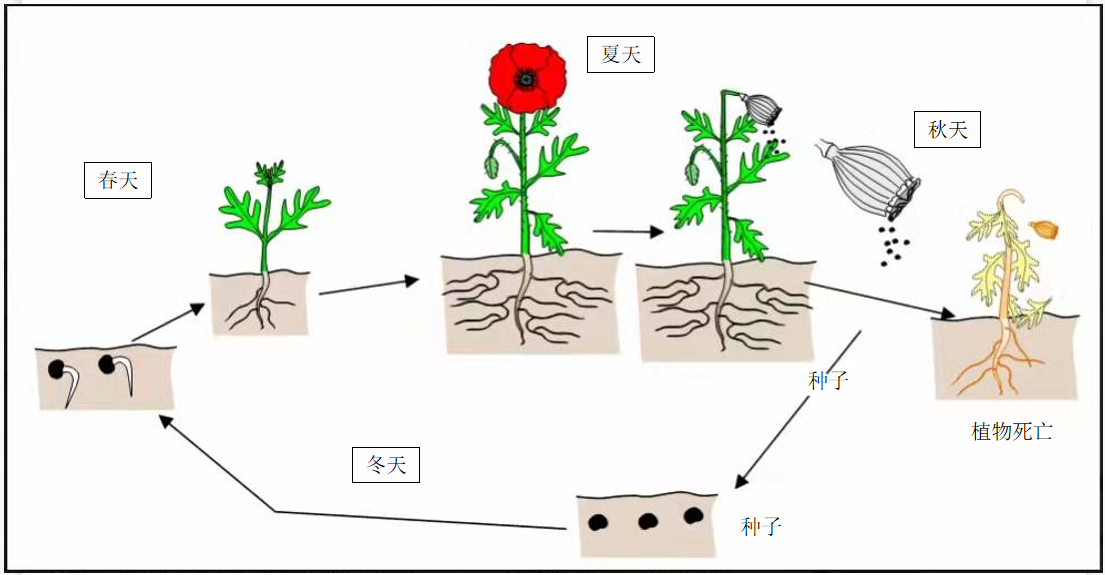
从这个角度来看,植物因为可以适应环境而显得格外引人注目。但它们很好地融入进人们的日常生活中以至于我们有时会忘记它们!它们就在那里…不变!在冬天,望着翻耕过的田野,能看见的就只有泥土,而到了春天,这里便被一抹熟悉的绿色覆盖着,于是,人们便意识到植物那非凡的生命力(图1,[1])。
无论风雨雪冻,亦或热浪向我们涌来…植物都在那里!能够适应高度波动的环境的确是它们的一大特性。植物必须根据昼夜和季节长短以及它们所处的环境来应对温度、光照和湿度的巨大差异。土壤性质也决定了植物生长发育的特殊条件,有时土壤中会存在矿物质营养(氮、磷等)严重不足的情况,相反,由于大量有毒金属(镉、铅、铝等)的存在,土壤中也会存在有害毒性物质。一些灌溉用水或者沿海陆地会引起盐胁迫(由土壤盐碱化引起的压力。这种盐碱化可以是自然形成的,也可以是由农业活动引起的,比如灌溉(使用低质量水)或者使用某些类型的化肥。这个地点具有相对均匀的物理和化学特性。这种环境是一系列生命形式的家园,包括植物群落、动物群和微生物。一个生物群落和它所支持的生物群落形成了一个给定的生态系统。有些植物喜欢阴暗的环境,比如蕨类植物,它们更喜欢在光线不足的地方生长,或者水生植物,比如水莼,它们需要大量的水。同样,钙质土壤会支持“在石灰岩上生长”的石灰植物。南法国的灌木丛植物就是这种情况。一些“逃离石灰岩”或者称为喜酸性土壤的植物,比如栗树或蕨类植物,则更喜欢在酸性土壤中生长。但植物不仅与它们的物理环境互动,它们还与其他生物体互动。一些生物可能对它们有益,比如通过促进它们的营养,例如共生细菌和菌根真菌。菌根是植物根系与土壤真菌之间的共生关系,影响超过95%的陆地植物。它们使植物更好地获取土壤养分,并帮助它们更好地抵抗环境压力。),从而干扰正常的植物营养过程。物理环境中的这些波动有利于植物依据自身对群落生境的适应能力进行地理分布。而其他能够感染它们的因素就对它们有害了,例如病毒、细菌、植物病原真菌,或者通过食用它们对其造成伤害,就像许多昆虫和食草动物那样。就像植物适应环境的物理变化一样,在进化的过程中,它们已经形成了抵御病原体侵袭的反应能力。
与动物不同,植物无法通过逃离来避免那些对其完整性和生存不利的或恶劣的环境条件。它们也没有动物那样的中枢神经系统,无法通过感官提供的信息而采取行动来适应变化的环境条件。根将它们固定在土壤中,并为地上部分提供水分和必需的矿物质元素:氮、磷、钾、硫、铁、锌、镁、锰……[3]。叶通过光合作用反应(光合作用是一种生物能量过程,它使植物、藻类和某些细菌能够利用阳光从大气中的二氧化碳合成有机物质。太阳能被用来氧化水并减少二氧化碳,从而合成有机物质(碳水化合物)。水的氧化导致大气中氧气的形成。光合作用是自养生物的基础,是叶绿体在细胞内的综合功能的结果。)[4](链接到文章“光合作用之光”)将太阳提供的光能转化为含碳有机分子(糖类、脂肪、蛋白质)。让我们简要回忆一下光合作用发生在特定的叶片上细胞器(在细胞内具有特定功能的特殊结构。例如,原子核、线粒体和叶绿体),叶绿体(光合真核生物细胞质的有机物细胞(植物、藻类)。叶绿体作为光合作用的场所,产生氧,在碳循环中起着至关重要的作用:它们利用光能固定二氧化碳并合成有机物。因此,它们负责植物的自养。叶绿体是大约15亿年前光合作用的原核生物(蓝藻类型)在真核细胞内共生的结果。)它们的叶绿素捕获太阳光子,导致水分子分裂以及氧气的释放,二氧化碳中的碳同化成有机分子,以及产生三磷酸腺苷(三磷酸腺苷的缩写。一个三磷酸核苷由腺嘌呤(氮基)、核糖(含5个碳原子的糖)和三个磷酸基团组成。一种既能提供能量又能储存能量的化合物,存在于所有生物体中。也用作核酸合成材料。)。
因此,植物叶片与外部环境之间的气体交换(水蒸气、氧气和二氧化碳)至关重要。从解剖学的角度来看,这些交换发生在非常特殊的叶片结构中:气孔(图 3 ;[5],[6]),它由两个表皮细胞组成,这种细胞称为保卫细胞。根据保卫细胞的膨胀程度(这是细胞伸长的状态,与植物或动物细胞中液泡或囊泡由于水进入而膨胀有关。),气孔选择打开或者关闭从而允许上述物质交换,这取决于温度、亮度或湿度等环境参数 [7]。因此,气孔功能和光合作用是帮助植物适应环境的两个重要因素,特别是对于干旱适应或生活在干旱环境如沙漠中的植物,稍后我们也会在文中对其进行讨论。
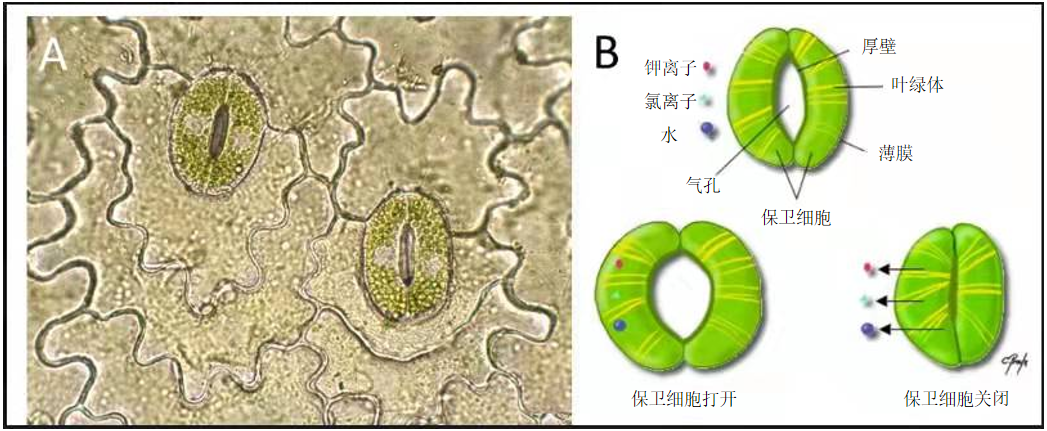
叶表皮的两个特殊细胞—保卫细胞,聚集形成一个气孔;[图片来源:photo © Christophe Charillon ;见参考资料 [5]
豆型的保卫细胞在其中间区域形成一个空的空间,即小孔,通过小孔,气体和水可以在叶子和外部环境之间进行交换。保护细胞膨胀与否意味其着开口可以打开或关闭。白天天气热时关闭气孔可以防止植物失水。气孔的关闭受离子流(钾、氯等)的影响以及邻近细胞间输水的影响。[图片来源:© Chantal Proulx; 见参考资料 [6]]
一方面,植物的生命是固定不动的,它们又缺乏感知外部环境的感觉器官,另一方面,它们也没有中枢神经系统,因此,植物已经进化到能够适应外界随着时间的推移而改变的环境条件。
植物适应的这两个方面并不使用相同的概念和机制。第一种情况是不同植物种对不同气候的空间适应。并不是所有的植物种都能随处生长,这种气候地理适应是基于有利性状的自然选择原则,这些性状随着时间的推移在物种的世代遗传中被继承下来(见“生物对其环境的适应”)。第二种情况是同一物种的植物对于特定地点波动环境条件的适应。因此,植物可以连续经历干旱、寒冷、强光等时期。它通过活跃生理过程来适应这些变化,通常是利用基因组表达的重塑从而达到较高的表型可塑性(在特定的环境条件下,生物体从给定的基因型中表达不同表型的能力)(见“适应:应对环境挑战”)。
因此,不同植物种基因组中固定的特征和它们基因组表达的可塑性相结合,使植物能够在不移动的情况下应对多种多样的环境条件。
2.植物的空间适应:从沙漠到针叶林
世界上不同经纬度以及不同海拔高度上都有植物的存在。然而,在不同的气候条件下,温度、光照和湿度状况千差万别。沙漠地区干燥缺水,热带雨林水分充足!观察生长于这些极端环境中的植物足以令人们发现它们通常不属于同一个物种并且具有反映所处生境特点的形态和结构特征[8]。
在此,我们提供两个截然不同的景观举例说明:针叶林(由冰水侵蚀形成的广阔湖泊系统穿越的森林型植被区,与亚北极气候密切相关,是主要的陆地生物群系之一。是针叶林和北极苔原之间的过渡带,位于西伯利亚地区。)和亚利桑那沙漠(图4)。

图4. 根据地理和气候的植被多样性。A. 针叶树(冷杉、云杉等)生长在西伯利亚的针叶林中,[来源:© Eniscuola.com] B.亚利桑那州的沙漠是巨型仙人掌和其他多肉植物的家园,[来源:© Derek Ostrovski]。
针叶林是北方森林(这是以针叶树为主并有一些硬木树的森林。在欧洲,主要由落叶树(如桦树)和/或针叶树组成,从波罗的海延伸至乌拉尔山脉。在加拿大,这是最大的植被区域,占据了该国55%的陆地面积。)和北极苔原(在寒冷气候地区的不连续植被形成中,包括一些草本植物、苔藓和地衣,甚至一些矮小的树木(如桦树)。苔原的特点是地面在一定深度处永久冻结(多年冻土)。它覆盖了北半球极地区的最北部,在55°至80°纬度之间,在裸露地面和冰雪之上。)之间的过渡地带。它包括灌木、针叶树以及桦树,并且它所覆盖的区域是连续而又开阔的。针叶林地处亚极地气候,这里夏季短暂凉爽,日照时间长,冬季则异常寒冷。最暖月均温在 10-15℃之间,但最冷月均温可能会降至-30℃以下。年降雨量通常不足 500 毫米。
沙漠是全球十分干旱的一个地区,这里的年降雨量不足200毫米,甚至经常只有100毫米,最显著的特点就是土地贫瘠,植物量稀少。这种缺水与年复一年的不规则降水密切相关。沙漠遍布全球,覆盖了大约三分之一的陆地面积,几乎是法国面积的100倍。它们主要分布在北回归线、西亚、澳大利亚内陆和极地地区。所有沙漠的共同特征就是缺水。这里的空气相对湿度较低(一般低于50%),以及晴朗的天空最能解释高温变化。在炎热的沙漠中,白天的温度能够达到50℃以上,紧接着晚上的温度又会下降至0℃以下。
针叶林中最常见的植物是松树、冷杉和云杉等针叶树种。大多数针叶树都是常绿的,因为它们的叶子(图5)不会随着温度下降而凋谢。树木不需要在来年春天消耗大量的能量重新长出叶子,因此,这一特征是一种非常重要的适应。在针叶林中,太阳总是处于地平线以下的位置,这里的土壤也就经常缺乏养分。这两个因素限制了树木可利用能量,而它们常绿这一情况又使它们将这些能量用于生长而不是生长叶片。除此之外,尽管这里的降水量很多,但地面数月冻结(见“永久冻土”)还是会阻止根系从中吸取水分。针叶树木的叶片是针状的而不是那种叶面积很大的,这使得它们在蒸腾的过程中可以减少水分丢失。另外,针叶含水量很少,从而降低了冰冻的风险。最后,针叶树的特殊习性也是一种显著的适应行为,从而避免积雪把树枝压断。

A.针叶树种的针型叶片在秋天不会凋落并且以此减小叶面积从而减少与外界的交换。[图片来源:© Margarethe Maillard, ENS Lyon]
B.仙人掌的叶子更小,可以简化为刺,从而减少水分流失。[图片来源:© Siquisai (CC-BY-SA-3.0) via Wikimedia Commons]
C.温带地区树木落叶形状的多样性。[图片来源:© Kaare Jensen (Harvard University), Maciej Zwieniecki (UC Davis)]
在沙漠中,没有针叶树!这里的植物区系非常独特(图4),它们对于干旱的环境条件有着很好的适应能力,水在这里是最珍贵的。这里是仙人掌等肉质植物(由于土壤和气候的特点而适应在干旱环境中生存的肉质植物(如油脂植物)的特征。)的领地,它们将叶面积降至最小,有时甚至简化为刺,然后通过茎来进行光合作用(图5)。生活在沙漠中的植物主要有三种适应方式:肉质化、耐旱和避旱。所谓肉质植物,包括仙人掌科的所有物种,它们具有能够在其幼叶、茎或根中储存水分的特性。它们必须要在短时间内大量吸水,因为沙漠中的降雨强度很低而且持续时间也不长,在强光照的影响下土壤也会迅速变干。几乎所有的肉质植物都有很长的根,这些根在土壤表层水平生长以更有效地捕获水源。一旦这些水分被吸收储藏,就不会丢失,这得益于沙漠植物缩小甚至消失的叶片。除此之外,这些叶片和茎上覆盖有一层蜡质层,这使它们几乎可以不受外界环境的影响。在生理水平上,许多肉质植物对水有一种非常有效的光合作用模式,称为“景天酸代谢”(CAM)的植物(见焦点“石莲花”)。CAM植物在夜间开放气孔以促进气体交换,在气孔关闭时储存白天光合作用需要的二氧化碳,从而减少水分丢失。由于夜间的低温和高湿度,CAM植物每单位碳水化合物损失的水分比白天气孔开放的植物(所谓的C3植物)少10%。
耐旱性是许多干旱地区植物的一种能力。这些植物能经受干旱(干燥的作用;使干燥:使植物的土壤干燥。除去物质中所含的天然或非天然的水分)却不死亡。通常,它们在干旱时期失去叶子并进入深度休眠状态。植物最大的水分流失方式就是叶片和气孔的蒸腾作用;因此,叶片的缺失有助于保存茎中的水分。有些植物没有这种弱化叶片的功能,它们的叶子上覆盖着一层减少水分蒸发的脂质。不像肉质植物那样只有表层的根系,一些沙漠树种和灌木能够存活下来,多亏了它们高度发达的根系,这些根系可以达到树冠(森林上层,直接受太阳辐射影响。被视为栖息地或生态系统,尤指在生物多样性特别丰富的热带森林中生物生产力。)表面的两倍,并且可以到达很深的地方。当强降雨来临的时候,深层土壤的保湿性会更好一些,从而给与这些植物更长的生长时间。另外,即使是在水很少或者没有水的情况下,这种类型的植物也能够保持光合活性,而水很少或者没有水对温带地区的绝大多数植物来说可以说是致命的。
在沙漠中发现的第三种植物……由于条件太差,大多数时间根本不存在。有些植物将其所有的能量都用于快速产生种子上来,而不是尽可能久地维持其营养状态,这种耐旱方式在一年生植物上是可行的。在许多沙漠中,由于降雨和气温的下降,秋季的气候条件通常来说是比较有利的。一年生植物的非休眠(生物体在生长发育较慢暂时停止的生命阶段的特性。)种子在这时可以快速大量萌发,并且在几周内完成它们的整个生命周期。然后它们就会产生足够多的种子,以确保该物种在冬季来临前的可持续性。
这两个特定植物群落在截然相反的环境(针叶林和沙漠)中适应生存的例子清楚地说明了自然选择在植物物种进化中的作用,这些植物物种很好地适应了特定的,甚至是极端的环境条件。随着时间的推移,使发育最适应外部条件的突变已被固定在物种的基因组中(例如叶形态,图5),从而确保该物种在特定生境中的可持续性。
3.植物的时间适应:风暴预警!
我们已经发现,在进化的压力下,植物种能够适应非常多样化的环境。然而,同属一个物种的个体表现出很高的可塑性,这使它们能够适应同一生境中波动着的环境条件[9]。最明显的一个例子就是生长在海边的树木,它们受到来自同一方向的强风影响,这对它们的结构产生了强烈的机械压力(图6)。

干旱适应的极端情况同样引人注目,比如说杰里科玫瑰 (Selaginella lepidophylla),通常被称为复活植物,它们看似已经死亡,但如果获得水就会很快复活(图7)。

这两个例子很好地说明了植物感知外界环境压力的能力,并对其作出反应从而尽可能地适应。植物对胁迫的感知以及由胁迫产生的生物反应是近二十年来人们努力研究的重点。分子遗传学(这包括对基因在分子水平上功能的分析)的发展及其与生物物理学、生物化学和生理学分析方法的结合,促进了综合性植物生物学的发展。这为了解胁迫感知和信号传递的机制提供了可能,而这种信号会促进基因表达的重新编程,最终表现为植物对胁迫的表型反应。
图8给出了植物适应特定胁迫条件(寒冷或高盐)的信号通路的两个例子。当寒冷发生时,植物体内会发生一系列反应并对寒冷反应的基因表达进行调节。首先,植物细胞质中的钙浓度增加,这使得许多修饰转录因子的酶被激活[10]。然后,这些因子会附着在冷响应基因或它们的调控因子的上游DNA上,从而激活或抑制它们的表达。当压力消失时,系统通过某些转录因子的其他后转录变化恢复平衡,从而使得转录因子被蛋白质水解(图8A)。
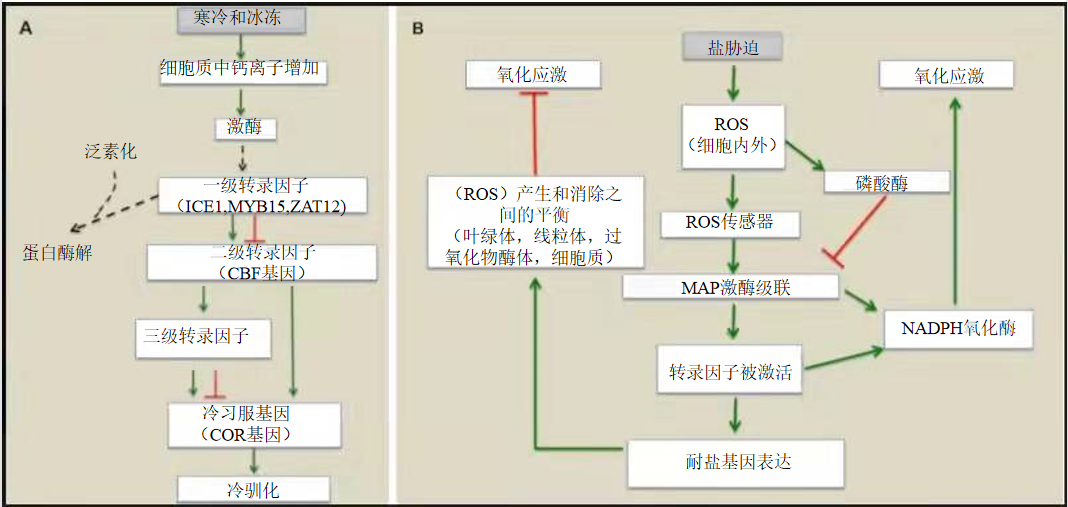
A.冷应激反映转录网络。冷诱导初级转录因子的激活,通过激酶的磷酸化在转录后对其进行修饰。一旦被激活,它们就会正向或负向地调节次级转录因子CBFs的表达。这些将激活三级转录因子的表达,并直接激活冷习服基因(COR基因)的表达。三级转录因子也正向或负向地调节COR基因的表达。当冷胁迫消失时,系统通过对初级转录因子泛素化的转录后修饰恢复平衡。这一反应使得初级转录因子进行蛋白水解,从而实现CBF基因表达的抑制。以横条结尾的红线表示负调控;以箭头结尾的绿线表示正调控。虚线表示转录后事件。
B.盐胁迫调节反映网络。以横条结尾的红线表示负调控;以箭头结尾的绿线表示正调控。ROS=活性氧;MAP=裂原活化蛋白。
植物对环境中高盐的反应是由过氧化氢(H2O2)、超氧离子(O2.-)或羟基自由基(-OH)等活性氧(ROS)的产生和消除之间的平衡决定的。植物传感器对这些活性氧的感知将会使得激酶被激活,激酶将转录因子磷酸化,从而激活它们。由这些转录因子调节的反应基因的产物将会导致活性氧的消失,从而消除过量盐引起的氧化应激。这一平衡还取决于以下条件:磷酸酶的激活将会平衡激酶的作用,而反过来,氧化酶的激活将促进氧化应激。
图8中所示方案存在许多变体反映了给定胁迫的特殊性,但目前被科学界广泛接受的一般原则有以下几点:
- 胁迫会使植物产生信号。
- 这些信号往往是来自代谢活动的有机小分子,它们可以是:
-植物细胞壁降解产生的多糖;
-由特定的酶产生的脂质分子,例如脂氧合酶—催化脂肪酸氧化的酶;
-植物伤流液中循环的小肽发出的长距离应激信号。
这些代谢物中有的能够发挥植物激素的作用,比如脱落酸,它被认为是一种真正的应激激素。 - 这些信号通常被位于细胞膜上的蛋白质受体所感知;它们具有激酶或磷酸酶活性,换而言之,它们能够去除或添加磷酸基团。
- 在许多情况下,信号的传播或放大需要次级信使的干预。钙离子和活性氧(ROS)是植物在应对环境变化时最常见的次级信使。
- 这些次级信使能够激活可溶性于细胞质和细胞核中的蛋白激酶和磷酸酶蛋白的级联反应。
- 最终,这些级联反应的末端靶点通常是转录因子,它们能够与应激反应基因的上游DNA相结合,并激活这些基因的表达。
- 这些基因的所有产物(结构蛋白、酶等)都允许植物对其所经历的逆境胁迫做出适应性的表型反应。
除了这种响应环境约束的基因表达调控外,还有表观遗传调控[11],[12](见“表观遗传学、基因组及其环境”和“表观遗传学:环境是如何影响我们的基因的”)。由于环境变化反复无常,对植物而言,拥有这些“过去的记忆”是有利的,它们可以利用这些储存信息更有效地适应新的变化。最著名的一个例子就是植物对食草动物的防御,但这些机制也与适应非生物胁迫有关。植物通过不同的方法生成这种记忆:代谢物的积累,比如说抗干旱的渗透调节物质,上述所提到的调节蛋白的磷酸化/去磷酸化。但很多研究都强调了表观遗传调控在植物适应不同胁迫方面的重要性,特别是叫做 miRNA 和 siRNA 的小调控 RNAs 发挥的作用。最初,有人描述了在适应低磷和低铜营养条件的情况下,植物适应环境限制的表观遗传调控。这些小 RNAs 在使植物适应干旱或温度升高方面的作用已经被阐明。在更完整的水平上,DNA 携带的基因在细胞核中被包裹在一个复杂的 DNA 和蛋白质组成的染色质。染色质的压缩状态决定了基因的表达。它受组蛋白的转录后修饰(甲基化、乙酰化、磷酸化等)调控,组蛋白是染色质内结构 DNA 的蛋白质(见“表观遗传学、基因组及其环境”)。因此,上述植物的逆境胁迫条件能够改变邻近基因的染色质结构,这些基因对于适应逆境胁迫至关重要。这一过程有助于调节胁迫基因的表达和植物的适应反应[13]。
4.展望气候变化背景下植物的适应性
目前,地球正处于气候变化之中,其表现为温度升高、降水模式改变以及大气二氧化碳浓度增加等等。众所周知,干旱和洪涝灾害会影响植物生命。因此,大气二氧化碳浓度增加作为决定性因素,使得植物在各种胁迫环境中必须不断强化自己的适应能力[14]。
一些研究分析了处于高浓度二氧化碳环境中不同物种的转录组(mRNA库,即基因表达)、蛋白质组(蛋白质库)和代谢组(代谢产物库)。人们观察到,在所有的这些水平上都发生了明显的重组,主要涉及光合作用和碳代谢,以及氨基酸、淀粉和糖类的生物合成。另一个受气候变化影响而显著改变的方面就是植物营养。温度和二氧化碳浓度的增加会影响土壤微生物的生理机能,从而改变土壤的养分循环及其对植物生长的有效性[15]。因此,在二氧化碳浓度等于2050年预期二氧化碳浓度的条件下下进行了植物生长实验。实验表明,在这种条件下,C3植物种子的铁、锌浓度明显下降。在高浓度二氧化碳环境下,C3植物的蛋白质含量也随氮的变化而降低。但是,CAM植物受二氧化碳增加的影响较小。
因此,这种气候演变将会改变某些物种的地理分布,并促进新的适应过程的出现,但也会通过改变植物的营养质量来影响人类活动[16]。这将对农业生产实践产生巨大影响。
参考资料和说明
封面图片。 [Source: Assignment http://www.ForestWander.com License; License CC-BY-SA 3.0]
[1] http://bip88.net/bip/index.php/activites-humaines/agriculture/40Semaine-9 & http://bip88.net/bip/index.php/activites-humaines/agriculture/40Semaine-2
[2] https://www.youtube.com/watch?v=9wLnavgmVjs
[3] https://fr.wikipedia.org/wiki/Nutrition_v%C3%A9g%C3%A9tale
[4] https://fr.wikipedia.org/wiki/Photosynth%C3%A8se
[5] http://acces.enslyon.fr/evolution/evolution/relations-de-parente/enseigner/activites-pratiques-et-classification/les-tp/tp-presence-de-stomates
[6] http://www.cours-pharmacie.com/biologie-vegetale/leau-de-labsorption-a-la-transpiration.html
[7] http://www.snv.jussieu.fr/bmedia/mouvements/nasties-stomate.htm
[8] http://www.mbgnet.net/bioplants/adapt.html
[9]http://www.bdesciences.com/nolaj/SVS/L3/Semestre%20VI/Replies%20of%20Plants%20a%20l%20Environment/Courses/Krys3000%20%282010%29/II%20-%20R%C3%A9response%20of%20plants%20to%20stress%20abiotics.pdf
[10] Transcription factor: protein necessary for initiating or regulating the transcription of DNA into RNA molecules.
[11] Tetsu Kinoshita T & Seki M (2014) Epigenetic Memory for Stress Response and Adaptation in Plants. Plant & Cell Physiology 55: 1859-1863. doi:10.1093/pcp/pcu125
[12] Sunkar R, Chinnusamy V, Zhu J & Zhu JK (2007) Small RNAs as bigplayers in plantabiotic stress responses and nutrient deprivation. Trends in Plant Science 12:301-309
[13] Kim JM, Sasaki T, Ueda M, Sako K & Seki M (2015) Chromatin changes in response to drought, salinity, heat, and cold stresses in plants. Frontiers in Plant Science 6:114. doi: 10.3389/fpls.2015.00114
[14] Ishita Ahuja, Ric CH de Vos, Atle M. Bonesand Robert & D. Hall (2010) Plant molecular stress responses face climate change. Trends in Plant Science 1:664-674. doi:10.1016/j.tplants.2010.08.002
[15] Pilbeam DJ (2015) Breeding crops for improved mineral nutrition under climate change conditions. Journal of Experimental Botany, 66:3511-3521. doi:10.1093/jxb/eru539
[16] Myers SS et al. (2014) Increasing CO2 threatens human nutrition. Nature 510:139-142. doi:10.1038/nature13179
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: BRIAT Jean-François (2024年3月14日), 植物固定的生命及其限制因素, 环境百科全书,咨询于 2026年7月29日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/fixed-life-of-plants-and-its-constraints/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。


